
Сетевидная и ганглионарная нервная система
Известно, что реактивность голодной амебы более высока, чем реактивность сытой амебы, и у нее создается повышенная готовность реагировать на соответствующие полезные воздействия или на раздражения, которые сигнализируют об их появлении.
Наконец, одноклеточное может "привыкать" к соответствующим воздействиям, снижая реакции на них по мере их длительного повторного предъявления.
Механизмы, лежащие в основе этого явления, еще мало изучены, и лишь проведенные в последнее время наблюдения заставляют думать, что появление и сохранение этих воздействий является результатом известных модификаций и рибонуклеиновой кислоты, составляющей один из основных компонентов их плазмы.
Следует отметить, что тело простейших является относительно однородным и процесс наиболее интенсивного обмена может возникать у него в месте непосредственно возникающего возбуждения, образуя тем самым временный "головной" конец его тела.
Наиболее сложные одноклеточные имеют несравненно более сложную структуру. У них можно различить постоянные "органы" в виде чувствительных участков протоплазмы, которые образуют, например, "жгутики" у бактерий. Эти "жгутики" находятся в постоянном движении и несут наиболее существенные функции ориентировки во внешней среде; в них и возникают градиенты постоянного возбуждения, которые, распространяясь но остальному телу одноклеточного, приводят его в движение.
Богатство поведения одноклеточных очень велико и подлежит еще специальному изучению, так же, как и те механизмы, которые лежат в его основе. Однако то, что мы уже знаем о них, заставляет думать, что здесь заложены основы той сложной деятельности активной ориентировки в среде, которые в дальнейшем составят важнейшую черту психической деятельности.
Происхождение нервной системы и ее простейшие формы
Описанные процессы раздражимости по отношению к биотическим воздействиям, чувствительности по отношению к нейтральным воздействиям, сигнализирующим о появлении жизненно важных воздействий, и элементарного сохранения следов достаточны для поддержания жизни одноклеточных животных.
Однако они становятся недостаточными с переходом к многоклеточным.
Переход к многоклеточным существенно усложняет условия существования. Питание путем прямой диффузии питательных веществ, занимавшее ведущее место на уровне простейших, здесь заменяется питанием дискретной (концентрированной) пищей; повышается роль активной ориентировки во внешней среде. Становится необходимым обеспечить гораздо более дифференцированные движения и гораздо более быструю проводимость возбуждения, чем та, которая была доступна путем постепенного распространения градиентов возбуждения по протоплазме одноклеточных. Это и приводит к значительному усложнению строения тела многоклеточного, к выделению клеток специализированной рецепции раздражений, доходящих до тела животного, и появлению первых сократительных клеток, несущих ту функцию, которую на дальнейших этапах эволюции возьмут на себя мышечные клетки. Это приводит, наконец, к тому, что в местах прежних градиентов возбуждения начинают откладываться дорожки наиболее возбудимой протоплазмы и образуют наиболее элементарную форму нервной системы, которая у этих животных носит характер диффузной, сетевидной нервной системы.
Все это с особенной отчетливостью можно видеть в строении тела относительно простых многоклеточных, относящихся к классу кишечнополостных, например в строении тела медузы, актинии или морской звезды.
Характерным для этой стадии развития многоклеточных является то, что проводимость возбуждения ускоряется благодаря появлению сетевидной нервной системы во много раз. Если проведение возбуждения по протоплазме не превышает скорости 1–2 микрон в секунду, то с появлением простейшей (сетевидной) нервной системы скорость проведения равна 0,5 метра в секунду (заметим, что при дальнейшем развитии нервной системы и переходе на следующие этапы ее усложнения скорость проведения возбуждения еще более возрастает, доходя у лягушки до 25 метров в секунду, а у высших позвоночных до 125 метров в секунду).
Однако значительные преимущества, которые возникают с явлением первичной диффузной (сетевидной) нервной системы, имеют и свои границы. Как и на описанных выше этапах эволюции, у животных с сетевидной нервной системой еще нет постоянного головного конца, управляющего их поведением. Возбуждение равномерно распространяется по сетевидной нервной системе на все тело животного, а место приложения внешнего раздражения становится временным ведущим пунктом. Только у наиболее сложных кишечнополостных одна часть тела (например, один луч у морской звезды), по своему строению не отличающаяся от других частей тела (лучей), может стать доминирующей, "ведущей" и берет на себя функцию наиболее активного органа при движении. Если у морской звезды отрезать (ампутировать) этот "ведущий" луч, то роль "ведущего" переходит к другому, расположенному рядом с ним лучу.
Естественно, что такое отсутствие постоянного ведущего органа, который мог бы не только воспринимать, но и перерабатывать, кодировать полученную информацию и создавать программы дифференцированного поведения, существенно ограничивает возможности поведения на этом уровне. Эти недостатки устраняются на дальнейших этапах эволюции, особенно с переходом к земному существованию и формированием более сложно построенной ганглионарной нервной системы.
Ганглионарная нервная система и появление простейших программ поведения
Переход к наземному существованию связан со значительным усложнением условий жизни. Прямая диффузия питательных веществ из окружающей среды становится невозможной, наличие готовой плотной (дискретной) пищи - несравненно меньшим. Эта пища находится теперь в резко неоднородной среде, и ориентировка, необходимая для получения пищи, значительно усложняется.
Все это создает необходимость дальнейшего усложнения организма животных, и прежде всего дальнейшую эволюцию сложных органов рецепции и движений и формирования сложных и централизованных аппаратов переработки информации и регулирования движений. Именно этому и соответствует следующий этап в эволюции нервной системы, приводящий к возникновению цепочечной, или ганглионарной, нервной системы, которая впервые появляется у червей и приобретает максимальную сложность у высших беспозвоночных, и прежде всего у насекомых.
Как появление ганглионарной нервной системы, так и формирование поведения, которое осуществляется с ее помощью, знаменуют важнейший скачок в эволюции жизнедеятельности.
Уже у наиболее простых беспозвоночных (червей) можно проследить совсем новый принцип организации нервной системы по сравнению с предыдущей стадией. На переднем головном конце червя сосредоточиваются волокна, которые кончаются химическими и тактильными рецепторами, расположенными особенно густо. Эти аппараты воспринимают химические, термические, световые изменения, происходящие во внешней среде, а также изменения влажности. Сигналы этих изменений проводятся по волокнам и доходят до переднего узла или ганглия, где они сосредоточиваются в нервном "центре", впервые появляющемся на этой ступени эволюции. Здесь эти сигналы перерабатываются (кодируются), и возникающие "программы" поведения в виде двигательных импульсов распространяются по цепочке нервных ганглиев, каждый из которых соответствует отдельному сегменту тела червя. Доходящие до этих ганглиев импульсы вызывают соответствующие движения, направление которых программируется и регулируется передним ганглием.
Здесь возникает новый принцип - централизованность нервной системы, резко отличающейся от принципа построения диффузной (сетевидной) нервной системы. Головной конец червя, где сосредоточена особенно густая сеть хемо-, механо-, термо-, фото - и гигрорецепторов, приобретает ведущую роль, в то время как сегментарные ганглии сохраняют лишь относительную автономию. Это легко проследить, если разрезать тело червя на две половинки. В этом случае передняя половина будет закапываться в землю, сохранив свои организованные движения, в то время как задняя половина будет лишь беспорядочно извиваться, не проявляя никаких признаков организованного движения.
Усложнение строения нервной системы на стадии червей позволяет проследить у них более совершенные (хотя еще очень примитивные) виды формирования новых, индивидуально приобретенных видов поведения. Это было показано в свое время известным американским психологом Р. Йерксом. Он помещал дождевых червей в Т - образную трубку, простейший лабиринт. В левом конце этой трубки червь получал электрический удар, вызывавший оборонительную реакцию. При многократном повторении этого эксперимента у дождевого червя можно было выработать "навык" избегать электрического удара и двигаться направо. Насколько медленно шел этот процесс, видно из следующего: понадобилось свыше 150 проб, чтобы поведение червя приобретало организованный характер и в подавляющем числе проб он начинал двигаться направо, избегая электрического шока (табл. 1.1).
Таблица 1.1 - Формирование новых видов поведения у дождевых червей
Ганглии нервной системы — скопление нейронов и глии, находящиеся за пределами головного и спинного мозга.
Подобные образования в центральной нервной системе называют ядрами. Они выступают связующими звеньями структур нервной системы, осуществляют первичную обработку импульсов, отвечают за функции висцеральных органов.
Человеческий организм осуществляет два типа функций – соматические и вегетативные. Соматические подразумевают восприятие внешних раздражителей и реакцию на них при помощи скелетной мускулатуры. Эти реакции могут контролироваться сознанием человека, а за их выполнение отвечает центральная нервная система.
Вегетативные функции – пищеварение, метаболизм, кроветворение, кровообращение, дыхание, потоотделение и другое, контролирует вегетативная система, которая не зависит от человеческого сознания. Кроме регуляции работы висцеральных органов, вегетативная система обеспечивает трофику мускулатуры и ЦНС.
Ганглии, отвечающие за соматические функции, представляют спинномозговые узлы и узлы черепных нервов. Вегетативные, в зависимости от расположения центров в ЦНС, делят на: парасимпатические и симпатические.
Первые находятся в стенках органа, а симпатические дистанционно – в структуре, называемой пограничным стволом.
Строение ганглия
В зависимости от морфологических особенностей, размер ганглиев колеблется от нескольких микрометров до нескольких сантиметров. По сути это скопление нервных и глиальных клеток, покрытое соединительной оболочкой.
Соединительнотканный элемент пронизан лимфатическими и кровеносными сосудами. Каждый нейроцит (или группа нейроцитов) окружен капсульной оболочкой, выстланной изнутри эндотелием, а снаружи волокнами соединительной ткани. Внутри капсулы располагается нервная клетка и глиальные структуры, которые обеспечивают жизнедеятельность нейрона.
От нейрона отходит один аксон, покрытый миелиновой оболочкой, который разветвляется на две части. Одна из них входит в состав периферического нерва и образует рецептор, а вторая направляется в ЦНС.
Вегетативные центры находятся в стволе головного мозга и спинном мозге. Парасимпатические центры локализуются в краниальном и сакральном отделах, а симпатические – в тораколюмбальном.
Ганглии вегетативной нервной системы
Симпатическая система включает два типа узлов: вертебральные и превертебральные.
Вертебральные располагаются с двух сторон от позвоночного столба, образуя пограничные стволы. Они связаны со спинным мозгом с помощью нервных волокон, которые дают начало белым и серым соединительным веткам. Выходящие из узла нервные волокна направляются к висцеральным органам.
Превертебральные расположены на большем расстоянии от позвоночника, при этом также дистанционно они находятся от органов, за которые они ответственны. Примером превертебральных узлов служат шейные, брыжеечные скопления нейронов, солнечное сплетение.
Парасимпаческий отдел образован ганглиями, расположенными на органах, или в непосредственной к ним близости.
Внутриорганные нервные сплетения размещены на органе или в его стенке. Крупные внутриорганные сплетения расположены в сердечной мышце, в мышечном слое стенки кишечника, в паренхиме железистых органов.
Ганглии вегетативной и центральной нервной системы имеют свойства:
- проведение сигнала в одну сторону;
- входящие в узел волокна перекрывают зоны влияния друг друга;
- пространственная суммация (сумма импульсов способна породить потенциал в нейроците);
- окклюзия (стимуляция нервов вызывает малый ответ, чем стимуляция каждого в отдельности).
Синоптическая задержка в вегетативных ганглиях больше, чем в аналогичных структурах ЦНС, а постсинаптический потенциал продолжительный. Волну возбуждения в ганглионарных нейроцитах сменяет депрессия. Эти факторы приводят к относительно низкому ритму импульсов, сравнительно с ЦНС.
Какие функции выполняют ганглии
Основным назначением вегетативных узлов является распределение и передача нервных импульсов, а также генерация локальных рефлексов. Каждый ганглий, зависимости от расположения и особенностей трофики, отвечает за функции определенного участка тела.
Ганглиям присуща автономность от ЦНС, что позволяет им регулировать деятельность органов без участия головного и спинного мозга.
В структуре внутристеночных узлов содержатся клетки – водители ритма, способные задавать периодичность сокращений гладкой мускулатуры кишечника.
Особенность связана с прерыванием, направляющимся к внутренним органам, волокон ЦНС на периферических узлах вегетативной системы, где они образуют синапсы. При этом непосредственное влияние на внутренний орган оказывают аксоны, выходящие из ганглия.
Каждое нервное волокно, приходящее в симпатический ганглий, осуществляет иннервацию до тридцати постганглионарных нейроцитов. Это дает возможность умножать сигнал и распространять импульс возбуждения, выходящий из нервного узла.
В парасимпатические узлах одно волокно осуществляет иннервацию не более четырех нейроцитов, а передача импульса осуществляется локально.
Ганглии – рефлекторные центры
Ганглии нервной системы принимают участие в рефлекторной дуге, что позволяет корректировать деятельность органов и тканей без участия мозга. В конце девятнадцатого века русский гистолог Догель в результате опытов по изучению нервных сплетений в желудочно-кишечном тракте, выявил три типа нейронов – моторные, вставочные и рецепторные, а также синапсы между ними.
Наличие рецепторных нервных клеток подтверждает и возможность трансплантации сердечной мышцы от донора реципиенту. Если бы регуляция сердечного ритма осуществлялась через ЦНС, после пересадки сердца нервные клетки подверглись дегенерации. Нейроны и синапсы в пересаженном органе продолжают функционировать, что свидетельствует об их автономности.
В конце двадцатого века экспериментальным путем установлены механизмы периферических рефлексов, которые делают превертебральные и интрамуральные вегетативные узлы. Возможность создавать рефлекторную дугу свойственна некоторым узлам.
Местные рефлексы позволяют разгрузить центральную нервную систему, делают регуляцию важных функций более надежность, способны продолжать автономную работу внутренних органов в случае прерывания связи с ЦНС.
Вегетативные узлы получают и обрабатывают информацию о работе органов, после чего направляют ее в мозг. Это вызывает рефлекторную дугу как в вегетативной, так и в соматической системе, что запускает не только рефлексы, но и сознательные поведенческие реакции.
65.Основной источник развития НС и его производные
Источник развития НС – нервная пластинка. Выделяясь из эктодермы, она последовательно превращается в нервный желобок (на этапе кот образуются нервные гребни) и нервную трубку. Из нервной трубки формируется головной и спинной мозг. Из нервных гребней – ПНС:СМУ, ганглии ВНС, эндокриноциты APUD- системы и хромаффинная ткань.
66. Уровни организации НС
1)клеточный ур-нь – нейроны, астроциты, олигодендроциты, эпендимоциты, микроглиоциты; 2) тканевой – нервная, астроглиальная, олигодендроглиальная, эпендимоглиальная и микроглиальная ткани.3) уровень структурно-функциональных единиц – паттерны, группы изоморфных и изофункциональных нейронов, модули, образованные из гетероморфных и гетерофункциональных нейронов, распределенные силы с-мы высших псих-х ф-й.
67.Нейрон – структурная и функциональная характеристика, онтогенез.
Нейрон – специализированная клетка, трансформирующая раздражение в нервный импульс, проводящая и передающая его, вызывающая адекватную деятельность эффекторного органа. Форма и величина зависят от числа отростков, длины аксона и кол-ва образуемых синапсов. Самые мелкие –короткоаксонные клетки (клетки-зерна коры мозжечка 5-6 мкм), самые крупные – длинноаксонные нейроны двигат центров.(100-120 мкм).В составе поверхности нейрона имеется 40% белков, 50% липидов, 10% углеводов. Гликокаликс нейрона толщиной 5-нм образован из полисахаридов, плазмолемма (10нм) состоит из двух электронно-плотных и светлого слоев, образованных из полярных гидрофильных и неполярных гидрофобных липидов. В них заключены молекулы интегральных и полуинтегральных белков. Структура: имеет ядро, цитоплазму и цитоплазматические органеллы: ЭПС, к Гольджи, митохондрии, лизосомы, микротрубочки и нейрофиламенты. Ядро – метаболический центр нейрона. В нем различают кариолемму, поры кариолеммы, кариоплазму, в кот погружены хромосомы, и ядрышко. Цитоплазматические структуры нейрона: а) в-во Ниссля, или базофильная субстанция, предоставляющая собой совокупность цистерн гранлярного эндоплазматического ретикулума; б) нейрофибриллярный аппарат, образующийся как артефакт в результате склеивания микротрубочек и микрофиламентов при импрегнации нервной ткани азотнокислым серебром .Онтогенез: нейроэпителиальная клетка(однослойная нервная трубка) – нейробласты (аполярный – биролярный – монополярный) – нейроны(монополярный – мультиполярный).
68Морфологическая и нейрохимическая классификация нейронов.
Морфологически нейроны классифицируются по числу имеющихся отростков. На этом основании различают: а)монополярный нейрон, обладающий однонаправленным проведением нервного импульса; б)униполярный нейрон, отросток которого проводит ретро- и анте-градные нервные импулься; в)биполярный нейрон, имеющий аксон и дендрит, г) псевдоуниполярный нейрон – видоизмененный биполяр; д) мультиполярный нейрон.
Нейрохимическая классификация основана на нейромедиаторе, с помощью которого передается нервный импульс. В соответствии с медиатором различают: а) холинергические нейроны, использующие ацетилхолин, б)моноаминергические нейроны, использующие норадреналин (норадренергические), дофамин (дофаминергические), адреналин(адренергические, гистамин(гистаменергическе, серотонин(серотонинергические; в)пептидергические нейроны, использующие для передачи импульса олигопептиды, н-р Y-пептид, VIP, в-во Р; г)пуринергические нейроны, тспользующие АТФ и ее производные.
69. Характеристика аксона и дендритов.Закон динамической поляризации нейрона.Дендриты – цитоплазматические отростки числом от одного до нескольких жесятков на нейроне. Общая площадь дендритов может превышать площадь тела нейрона на 1-2 порядка. В дендритах имеется базофильное в-во, короткие микротрубочки, митохондрии, элементы эндоплазматического ретикулума. Самой характерной структурой дендрита явл шипиковый аппарат. Каждый шипик – это рецептивная площадка, воспринимающая нервный импульс. Аксон – единственный длинный отросток, проводящий и передающий нервный импульс на исполнительный орган, составляющий 99% всей массы нейрона. Имеет веретеновидный формы утолщения, где происходит синтез, накопление и выделение нейромедиатора. По аксону совершается антеградный (200-400мм/сутки) и ретроградный (100-200мм/сутки) аксональный сок.Закон сформулировал Кахаль. Импульс передается от дендритов к ткани, далее к аксонам и только в такой последовательности.
70. Глия, ее разновидности и функции.
Глия – это вторичный элемент мозга, определенная среда, в которой существуют и функционируют нейроны. По строению и ф-и, а также по источникам развития нейроглию делят на макроглию( астроглия, олигодендроглия и эпендимная глия) и микроглию (мезоглию) Ф-и: опорно-механическая, изолирующая, трофическая, секреторная, защитная.
71. Нейронная теория – сущность и доказательства
Согласно нейронной теории НС имеет клеточную организацию.Нейронная теория доказывает:1) морфологическую самостоятельность нейронов и расчлененность НС, 2) интегрированность нейронов в морфофункциональные единицы: паттерны, модули, распределенные с-мы;3) специфическую нейрохимическую организацию нейронов и взаимод-е их между собой с помощью медиаторов или электрически; 4) многоуровневую иерархическую организацию НС, соподчиненность нейронов рабочих центров командным нейронам. Док-ва: а) наличие между нейронами синапсов, б) их вторичное образование при формировании НС, в) дегенерацию нейрона до ближайшего синапса.
72. Определение синапса, классификация синапсов, понятие об эфапсах и аутопсах
Синапс – зона пограничного раздела последовательно сочетающихся нейронов, где нервный импульс претерпевает количественные и качественные изменения. В каждом синапсе различают: а)пресинаптическую часть, содержащую синаптические везикулы с нейромедиатором;б) постсинаптическую часть, имеющую специфические рецепторы к медиатору; синаптическую щель шириной 15-30 нм. По морфофункциональным признакам синапсы делятся на химические (пузырчатые) и электрические, характеризующиеся плотным прилежанием плазмолеммы двух нейроцитов (преимущественно их дендритов или перикарионов). Химические синапсы могут быть возбуждающие и тормозные. Для них характерно присутствие в пресинаптическом полюсе пресинаптических пузырьков, заполненных медиатором — веществом, участвующим в передаче возбуждения на постсинаптический полюс. Наиболее распространенными миедиаторами являются норадреналин (в адренергических синапсах) и ацетилхолин (в холинергических). В химическом синапсе различают пресинаптическую часть, постсинаптическую часть и синаптическую щель. В пресинаптической части присутствуют пресинаптические пузырьки. В холинергических синапсах они светлые, в адренергических имеют плотную сердцевину и называются плотными пузырьками. Несвободные инкапсулированные окончания помимо терминалей нервного волокна и глиального аппарата имеют еще и соединительно-тканную капсулу. Межнейронные синапсы — специализированные контакты нервных клеток, проводящие импульсы в одном направлении. По морфологическим признакам среди них различают: 1. аксосоматические синапсы (терминальные ветви аксона первого нейрона заканчиваются на теле второго), 2. аксодендритические синапсы (терминальные ветви аксона первого нейрона вступают в синаптическую связь с дендритом второго), 3. аксоаксональные синапсы. Эфапс – электрический синапс. Различают:эфапс 1 типа-пропускает ток в одном направлении, эфапс 2 типа –пропускает ток в обоих направлениях. Аутопс – это контакт, который образован реккурентной ветвью аксона на теле или дендрите собственного нейрона.Принципы объемной трансмисси: передача нервного импульса при отсутствии сформированных синапсов одновременно на большие группы клеток. Полностью принципами объемной трансмиссии обладает молекула оксида азота с нитрооксидергическимим нейронами.
74. Гистогенез нервной системы и развитие нейронов.
1)система одиночных нейронов (гидроидные полипы), 2)сетевидная нервная система (медузы), 3) ганглионарная с-ма (большинство первичноротых), 4) цереброспинальная система позвоночных.Миоэпителиальная клетка –секреторный нейродоцит (губки), нейросекреторный униполяр (плпнария); нейросекреторно-трансмиттерный униполяр(двучтворчатые моллюски) –трансмиттерный униполяр (животные с ганглионарной НС) – мультиполярный нейрон (позвоночные).
75. СМУ и первичночувствительные нейроны, их классификация, величина и значение, нейрохимическая специализация.
Спино-мозговые узлы развиваются из нервных гребней. Ганглий покрыт капсулой, его интерстиций образует соединительную ткань и кровеносные каппиляры. В интерстиции расположены первичночувствительные нейроны (протонейроны), вокруг которых лежат сателлиты – клетки периферической олигодендрологии. Протонейрон – ПУК, дендрит которой образует на периферии нервные рецепторы, а аксон в составе задних корешков идет в спинной мозг. По величине тела различают: а)малые С-нейроны (15-30 мкм) –воспринимающие, передающие и дискринимирующие боль, образуют температурные и болевые рецепторы; б) В-нейроны(31-60мкм) – воспринимают тактильную, частично проприоцептивную чувствительность, осязание и давление, в) А-нейроны (61-120 мкм) – иннервируют мышцы, суставы и сухожилия. Большинство А и В нейронов для передачи нервного импульса используют глутамат, а С-нейроны – аспартат и в-во Р. Функции: Протонейрон - это первый(чувствительный) нейрон рефлекторной дуги соматической и вегетативной НС, это первый нейрон всех восходящих центрипетальных проводящих путей. Нейрон, поддерживающий трофику тканей. При его удалении или повреждении развиваются тяжелые дистрофические изменения, известные как деафферентационный феномен.
76. Пластинчатая и ядерная организация серого в-ва спинного мозга. Понятие о клеточной колонке.
Серое вещество занимает промежуточную часть спинного мозга – массивное серое в-во с передними, задними рогами и промежуточной частью с телами нейронов, преимущественно безмякотными волокнами. 1-3 и частично 4 пластины заняты нейронами студневидного вещества заднего рога. В 6 и частично 7 пластинах располагаются базальные ядра, обрабатывающие основной материал, поступающий из головного мозга. В 7 пластине находится ядро бокового рога, интерстициальное ядро кахаля, переключающее поток информации с протонейрона на мотонейрон.В 9 пластине находятся мотонейроны, собранные в ядра (латеральное –пеерднее и заднее, медиальное – переднее и заднее, центральное). В 10 пластине нейроны локализованы около центрального канала. Ядерная организация по Кахалю: в заднем роге аксоны протонейронов( 1 краевая зона Лиссоуэра), далее ворота Боля(2 роландовое вещество), крупные мультиполярные нейроны образут 3 собственное ядро заднего рога. Аксоны формируют спиноталамический путь общей чувствительности боковых канатиков. В основании заднего рога мультиполярные нейроны образуют 4 грудное ядро Кларка. Аксоны формируют спинномозжечковый путь Флекселя. Промежуточная зона: ядро Кахаля, два базилярных ядра – ничходящие пирамидные и экстропирамидные пути, латеральное ядро, медиальное ядро (спинномозжечковый путь Говерса), передние рога: два латеральных ядра – сгибатели, разгибатели конечностей, два медиальных – сгиб/разгиб туловища, центральное ядро – сгиб/разгиб плечевого и тазового пояса.Клеточная колонка - нервные клетки в сером в-ве собраны в колонки в сагиттальном разрезе. Каждая колонка иннервирует определенную группу мышц, а каждая клетка- строго определенную группу волокон в каждой мышце (двигательная единица), т.е. спинной и головной мозг имеют соматотопическую организацию.
77. Основные типы нейронов спинного мозга и центры ноцицептивной, висцеральной и проприоцептивной чувствительности. Роландово вещество как нервный центр боли
Различают три основных типа: 1)идиодендритический нейрон – снабжен большим числом дендроцитов; характерен для передних рогов, ядра Кларка и студневидного вещества заднего рога; 2)аллодендритический нейрон – с умеренно развитой дендрической короной; характерен для дорзальной части передних и вентральной части задних рогов; 3) изодендритический, или ретикулярный, нейрон- филогенетически более древний, с небольшим числом дендритов, характерен для промежуточной зоны. Экстероцептивная чувствительность- болевая, температурная и тактильная – ориентирована на нейроны студневидного в-ва и собственного ядра заднего рога; висцеральная чувствительность – преимущественно на нейроны промежуточной зоны и проприоцептивная – на ядро Кларка, тонкое и клиновидные ядра.
78. Нервный аппарат спинного мозга – собственный и координационный, их организация.
Центральная нервная система человека осуществляет контроль над деятельностью его организма и разделяется на несколько отделов. Головной мозг посылает и получает сигналы из организма и после их обработки имеет информацию о процессах. Нервная система разделяется на вегетативную и соматическую нервную системы.
Отличия вегетативной и соматической нервной системы
Соматическая нервная система регулируется сознанием человека и может управлять деятельностью скелетной мускулатуры. Все компоненты реакции человека на внешние факторы находятся под контролем полушарий мозга. Она обеспечивает сенсорные и моторные реакции человека, контролируя их возбуждение и торможение.
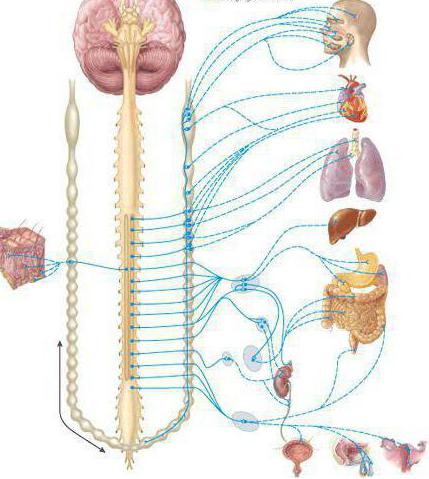
Вегетативная нервная система контролирует периферическую деятельность организма и не контролируется сознанием. Для нее характерны автономность и генерализованность воздействия на организм при полном отсутствии сознания. Эфферентная иннервация внутренних органов позволяет ей контролировать обменные процессы в организме и осуществлять обеспечение трофических процессов скелетной мускулатуры, рецепторов, кожи и внутренних органов.
Строение вегетативной системы
Работа вегетативной нервной системы контролируется гипоталамусом, который находится в центральной нервной системе. Вегетативная нервная система имеет метасегментарное строение. Ее центры находятся в головном, спинном мозге и коре головного мозга. Периферические отделы образованы стволами, ганглиями, сплетениями.
В вегетативной нервной системе различают:
- Симпатическую. Ее центр расположен в грудопоясничном отделе спинного мозга. Для нее характерны паравертебральные и предвертебральные ганглии ВНС.
- Парасимпатическую. Ее центры сосредоточены в среднем и продолговатом мозге, крестцовом отделе спинного мозга. Нервные узлы в основном интрамуральные.
- Метасимпатическую. Иннервирует желудочно-кишечный тракт, сосуды, внутренние органы организма.
В состав ее входит:
- Ядра нервных центров, расположенных в головном и спинном мозге.
- Вегетативные ганглии, которые расположены по периферии.
- Нервные волокна.
Рефлекторная дуга автономной нервной системы
Рефлекторная дуга вегетативной нервной системы состоит из трех звеньев:
- чувствительное или афферентное;
- вставочное или ассоциативное;
- эффекторное.

Чувствительное звено
Чувствительное звено расположено в спинномозговом ганглии. Этот ганглий имеет нервные клетки, сформированные группами, и их контроль осуществляется ядрами центрального головного мозга, большими полушариями и их структурами.
Чувствительное звено представлено частично униполярными клетками, которые имеют один приносящий или уносящий аксон, и они принадлежат спинальным или черепно-мозговым узлам. А также узлами блуждающих нервов, имеющих строение, похожее на спинальные клетки. В это звено входят клетки Догеля II типа, которые являются компонентами вегетативных ганглиев.
Вставочное звено
Вставочное звено в вегетативной нервной системе служит для передачи через низшие нервные центры, которыми являются вегетативные ганглии, и осуществляется это через синапсы. Расположено оно в боковых рогах спинного мозга. Нет прямой связи от афферентного звена на преганглионарные нейроны для их связи, существует кратчайший путь от афферентного нейрона до ассоциативного и от него до преганглионарного нейрона. Передача сигналов и нервных импульсов от афферентных нейронов в различных центрах осуществляется с различным количеством вставочных нейронов.
Например, в дуге спинального автономного рефлекса между чувствительным и эффекторным звеном существует три синапса, два из которых расположены в спинном мозге, а один в вегетативном узле, в котором расположен эфферентный нейрон.
Эфферентное звено
Эфферентное звено представлено эффекторными нейронами, которые расположены в вегетативных узлах. Их аксоны образуют безмиелиновые волокна, которые в составе со смешанными нервными волокнами иннервируют внутренние органы.
Вегетативные рефлекторные дуги расположены в боковых рогах.
Строение нервного узла
Ганглий – это скопление нервных клеток, которые имеют вид узелковых расширений толщиной около 10 мм. По своему строению вегетативный ганглий сверху покрыт соединительнотканной капсулой, которая образует строму из рыхлой соединительной ткани внутри органов. Мультиполярные нейроны, которые строятся из округлого ядра и крупных ядрышек, состоят из одного эфферентного нейрона и нескольких расходящихся афферентных нейронов. Эти клетки относятся по своему типу к клеткам головного мозга и являются двигательными. Их окружает неплотная оболочка – мантийная глия, которая создает постоянную среду для нервной ткани и обеспечивает полноценное функционирование нервных клеток.
Вегетативный ганглий имеет диффузное расположение нервных клеток и множество отростков, дендритов и аксонов.
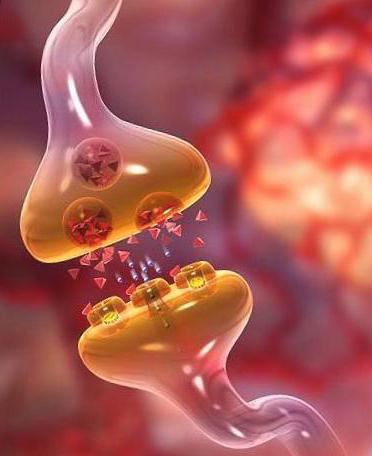
Спинномозговой ганглий имеет нервные клетки, которые расположены группами, и их расположение имеет порядок обусловленный.
Вегетативные нервные ганглии разделяются на:
- Сенсорные нейроны, которые расположены близко к спинному или центральному отделу мозга. Униполярные нейроны, из которых состоит этот ганглий, представляют собой приносящий или уносящий отросток. Они служат для афферентной передачи импульсов, и их нейроны образуют бифуркацию при ветвлении отростков. Эти отростки передают информацию от периферии к центральному афферентному нейрону – это периферический отросток, центральный – от тела нейрона в мозговой центр.
- Моторные, двигательные состоят из эфферентных нейронов, и в зависимости от их положения их называют паравертебральные, предвертебральные.
Симпатические ганглии
Паравертебральные цепочки ганглиев расположены вдоль позвоночного столба в симпатических стволах, которые идут длинной вереницей от основания черепа до копчика.
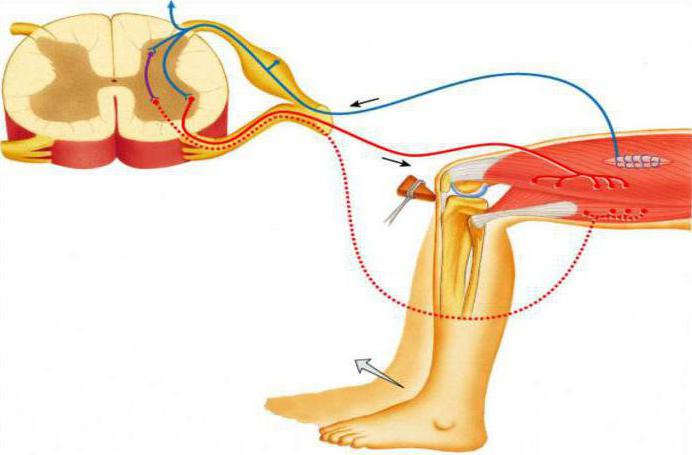
Предвертебральные нервные сплетения находятся ближе к внутренним органам, и их локализация сосредоточена перед аортой. Они образуют брюшное сплетение, которое состоит из солнечного, нижне- и верхнебрыжеечного сплетений. Они представлены двигательными адренергическими и тормозящими действие холинергическими нейронами. Также связь между нейронами осуществляется преганглионарными и постганглионарными нейронами, которые используют медиаторы ацетилхолин и норадреналин.
Интрамуральные нервные узлы имеют нейроны трех типов. Их описание было сделано русским ученым Догелем А.С., который, исследуя гистологию нейронов вегетативной нервной системы, выделил такие нейроны, как длинноаксонные эфферентные клетки первого типа, равноотросчатые афферентные клетки второго типа и ассоциативные клетки третьего типа.
Рецепторы ганглиев
Афферентные нейроны выполняют узкоспециализированую функцию, и их роль заключается в восприятии раздражителей. Такими рецепторами являются механорецепторы (реакция на растяжение или давление), фоторецепторы, терморецепторы, хеморецепторы (отвечают за реакции в организме, химические связи), ноцицепторы (реакция организма на болевые раздражители – это повреждение кожи и другие).
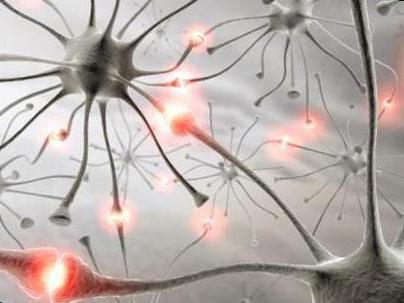
В симпатических стволах эти рецепторы по рефлекторной дуге передают в центральную нервную систему информацию, которая служит сигналом о повреждениях или нарушениях в организме, а также нормальной его работе.
Функции ганглиев
Каждый ганглий имеет свое местоположение, кровоснабжение, и его функции определяются этими параметрами. Спинномозговой ганглий, имеющий иннервацию из ядер головного мозга, обеспечивает непосредственную связь процессов в организме через рефлекторную дугу. От этих структурных компонентов спинного мозга иннервируются железы, гладкая мускулатура мышц внутренних органов. Сигналы, поступающие по рефлекторной дуге, идут медленнее, чем в ЦНС, и они полностью регулируются вегетативной системой, также она обладает трофической, сосудодвигательной функцией.
Читайте также:
