
Синапс это результат созревания нервных клеток
1. Проклассифицируйте нейроны по функциональному назначению. Каково функциональное назначение мотонейронов и каковы последствия их повреждения?
Нейроны делятся на три группы: афферентные (чувствительные), воспринимающие информацию о внешней или внутренней среде и передающие ее в ЦНС; эфферентные (двигательные, или мотонейроны), передающие информацию из ЦНС к органам-эффекторам и вставочные, обеспечивающие связь между афферентными и эфферентными нейронами. Мотонейроны – специализированные нейроны, которые передают двигательную информацию. В результате их повреждения развиваются параличи: утрата произвольных движений, повышение или снижение мышечного тонуса, повышение сухожильных рефлексов, утрата рефлекторных движений, атрофия пораженных мышц и др.
2. Каковы особенности созревания нейронов в онтогенезе.
Число нейронов в ЦНС становится максимальным уже на 24-ой неделе внутриутробного развития и больше не увеличивается. Растут лишь тела нейронов, увеличивается длина и количество их отростков. Детские нейроны чувствительны к недостатку кислорода и действию токсинов. У детей раннего возраста нейроны обладают низкой возбудимостью и лабильностью, поэтому легко подвержены запредельному торможению. В норме они должны переходить из состояния бодрствования в сон и, наоборот, в соответствии с циркадианными ритмами.
3.Приведите основные классификации синапсов: А –по веществу, –нейромедиатору, выделяющемуся в синапсах, Б– по характеру воздействияна нервную систему.
Синапсы – контакты между нейронами. По нейромедиатору синапсы подразделяютсяна:адренэргические, холинэргические, серотонинэргические, дофаминэргические, глицинэргические и др.По характеру воздействия на нервную систему синапсы бывают возбуждающие и тормозные.
4. Опишите факторы, необходимые для созревания межнейронных синапсов.
Синапсы активно формируются с момента рождения до 3 лет. Больше синаптических контактов разовьется в том случае, если ребенок получит из внешней среды множество сенсорных стимулов (демонстрация ярких игрушек, мультфильмов, чтение сказок и др.). Обучениеребенка различным видам деятельности способствует росту и ветвлению аксонов, образованию на их окончаниях синапсов в головном мозге. Образовавшиеся новые межнейронные связи способствуют повышению возможностей мозга к запоминанию и мышлению.
5Что такое миелинизация нервных волокон? Какая часть двигательных нервных волокон миелинизируется после рождения?
Созревание нейронов сопровождается процессом миелинизации – формированием вокруг нервного волокна защитной миелиновой оболочки, способствующей точному и изолированному проведению нервного импульса. После рождения миелинизируется 75 % нервных волокон. Последствия повреждения миелиновой оболочки: замедление или блокада проведения нервных импульсов.
6. Каковы особенности проведения возбуждения по нервному волокну ребенка по сравнению с проведением возбуждения у взрослого?
У детей раннего возраста проведение возбуждения более медленное и не полностью изолированное из-за меньшего диаметра нервных волокон и неполной их миелинизации. К 10–12 годам у детей заканчивается созревание нервов и скорость проведения возбуждения по ним становится, как у взрослых. Соответственно, по мере взросления, совершенствуются и рефлекторные реакции.Физиологические свойства мякотных нервных волокон: возбудимость, проводимость и лабильность; величина потенциала покоя и потенциала действия. В процессе созревания волокон эти свойства увеличиваются.
7.К какому возрасту формируются проводящие нервные пути двигательные и чувствительные, а также ассоциативные нервные связи в коре головного мозга?
Развитие и миелинизация двигательных путей, берущих начало в коре больших полушарий заканчивается к 14 годам. Развитие чувствительных путей (спинно-мозжечковых и спинно-таламических) завершается к 7–14 годам. К 17–18 годам завершается миелинизация ассоциативных волокон, обеспечивающих связи разных долей и зон коры. Развитие проводящих двигательных и чувствительных путей способствует активному совершенствованию координации движений, вовлечению подростков в хореографию, спортивно-оздоровительную работу, укреплению физического здоровья. Благодаря развитию ассоциативных нервных связей в коре высокого уровня развития достигает нервно-психическая и познавательная деятельность.

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.
I
Синапс (греч. synapsis соприкосновение, соединение)
специализированная зона контакта между отростками нервных клеток и другими возбудимыми и невозбудимыми клетками, обеспечивающая передачу информационного сигнала. Морфологически С. образован контактирующими мембранами двух клеток. Мембрана, принадлежащая отросткам нервных клеток, называется пресинаптической, мембрана клетки, к которой передается сигнал, — постсинаптической. В соответствии с принадлежностью постсинаптической мембраны С. подразделяют на нейросекреторные, нейромышечные и межнейрональные. Последние в зависимости от места их расположения разделяют на аксодендритические, аксосоматические, аксо-аксональные и дендро-дендритические. Наиболее сложно устроено нервно-мышечное соединение, называемое двигательной концевой пластинкой, в котором окончание аксона образует множественные синаптические контакты со специализированной мышечной мембраной. Отдельные нейроны могут иметь множество синаптических контактов с другими клетками (рис. 1).
По способу передачи возбуждения с пресинаптической на постсинаптическую мембрану выделяют химические и электрические (так называемые эфапсы) синапсы. В синапсах с химической передачей возбуждения между пре- и постсинаптической мембранами имеется синаптическая щель, куда выделяется химическое вещество-передатчик — медиатор (Медиаторы). Химические С. часто обозначают по названию медиатора (например, холинергические, адренергические, серотонинергические и т.п.). В эфапсе пре- и постсинаптические мембраны плотно соприкасаются и возбуждение передается посредством электрического тока. В зависимости от изменения биопотенциала постсинаптической мембраны различают С. деполяризующие, или возбуждающие, и гиперполяризующие, или тормозные.
Механизм передачи возбуждения принципиально одинаков во всех химических С. В нем можно выделить следующие основные этапы: синтез и депонирование медиатора в пресинаптическом нейроне и его окончаниях; высвобождение медиатора из депонирующих везикул и его выход в синаптическую щель; взаимодействие медиатора со специфическими хеморецепторами постсинаптической мембраны с последующей генерацией биоэлектрического потенциала; инактивация выделенного медиатора с помощью ферментов или системы обратного поглощения.
Биологически активные вещества, выполняющие функцию медиаторов, делят на несколько групп. К классическим нейромедиаторам относят ацетилхолин, адреналин и норадреналин, дофамин, серотонин, а также аминокислоты глицин и глутаминовую, аспарагиновую и гамма-аминомасляную (ГАМК) кислоты. Отдельно выделяют нейропептиды: вещество П, энкефалин, соматостатин и др. Медиаторную роль могут выполнять также АТФ, гистамин, пуриновые нуклеотиды. В соответствии с принципом Дейла, каждый отдельный нейрон в своих синаптических окончаниях один и тот же медиатор, поэтому нейроны можно обозначать также по виду медиатора: холинергические, адренергические, ГАМК-эргические, пептидергические (см. Нейросекреции (Нейросекреция)). Синтез нейромедиаторов осуществляется как в соме нейрона с последующим аксонным транспортом, так и непосредственно в пресинаптических окончаниях аксона, где медиатор концентрируется в везикулах, или синаптических пузырьках.
Основные физиологические свойства С. обусловлены механизмом передачи возбуждения (Возбуждение). Наличие пресинаптической мембраны с медиатором и хеморецепторов на постсинаптической мембране обеспечивает одностороннюю передачу возбуждения. Время освобождения медиатора из везикул при экзоцитозе, диффузия медиатора через синаптическую щель, взаимодействие медиатора с клеточными рецепторами постсинаптической мембраны и формирование потенциала действия создают так называемую синаптическую задержку в передаче возбуждения через С. Ее продолжительность для теплокровных животных составляет 0,2—0,5 мс. Величина синаптической задержки указывает на низкую лабильность С. по сравнению с нервными волокнами и мышцами. В связи с этим С. легко утомляется. Наличие специфических хеморецептивных участков на постсинаптической мембране делает С. высокочувствительным к биологически активным веществам. Хеморецептивные зоны часто являются точкой приложения как лекарственных средств, так и различных токсических веществ.
Библиогр.: Боголепов Н.Н. Ультраструктура синапсов в норме и патологии, М., 1975; Физиология человека, под ред. Р. Шмидта и Г. Тевса, пер. с агнл., т. 1, с. 78, М., 1985; Экклс Дж. Физиология синапсов, пер. с англ., М., 1966.
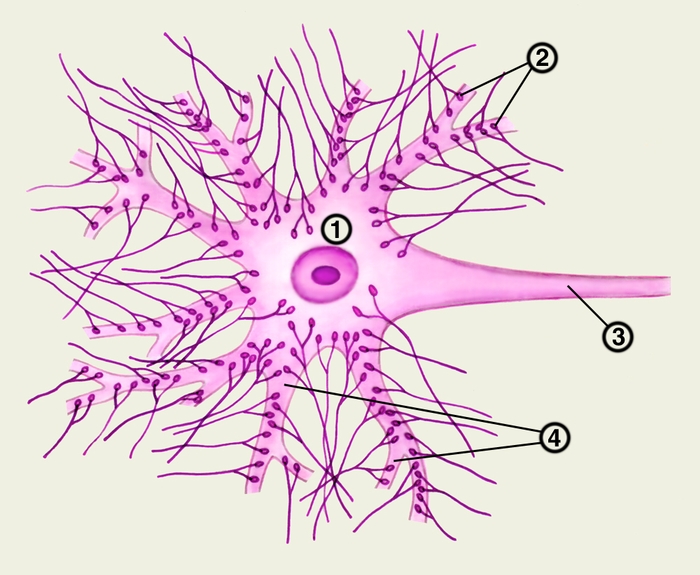
Рис. 1. Синаптические контакты нейрона: синаптические бляшки (2) окончаний пресинаптических аксонов образуют соединения на дендритах (4) и соме (1) постсинаптического нейрона; 3 — аксон нейрона.
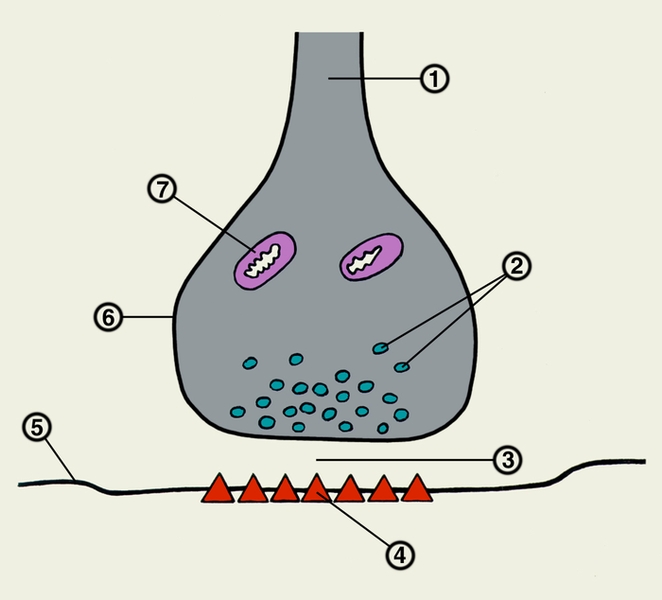
Рис. 2. Межнейрональный синапс: 1 — нервное волокно (аксон); 2 — синаптические пузырьки; 3 — синаптическая щель; 4 — хеморецепторы постсинаптической мембраны; 5 — постсинаптическая мембрана; 6 — синаптическая бляшка; 7 — митохондрия.
II
специализированная структура, обеспечивающая передачу нервного импульса с нервного волокна на какую-либо клетку или мышечное волокно, а также с рецепторной клетки на нервное волокно.
Синапс адренергический (s. adrenergica) — С., в котором медиатором является норадреналин.
Синапс аксо-аксональный (s. ахо-axonalis, LNH) — С. между двумя аксонами.
Синапс аксодендритический (s. axodendritica, LNH) — С. между аксоном одного нейрона и дендритом другого.
Синапс аксомышечный (s. axomuscularis, LNH) — см. Синапс нервно-мышечный.
Синапс аксосоматический (s. axosomatica, LNH) — С. между аксоном одного нейрона и телом другого.
Синапс аксоэпителиальный (s. axoepithelialis, LNH; син. С. нейросекреторный) — С. между секреторным нервным волокном и гландулоцитом.
Синапс возбуждающий — С., в котором в результате деполяризации постсинаптической мембраны генерируется возбуждающий постсинаптический потенциал.
Синапс дендро-дендритический (s. dendrodendritica, LNH) — С. между дендритами двух или нескольких нейронов.
Синапс межнейронный (s. Interneuronalis, LNH) — общее название С. между какими-либо элементами двух нейронов.
Синапс нейромускулярный — см. Синапс нервно-мышечный.
Синапс нейросекреторный (s. neurosecretoria) — см. Синапс аксоэпителиальный.
Синапс нервно-мышечный (s. neuromuscularis; син.: аппарат нервно-мышечный, бляшка двигательная — устар., бляшка моторная — устар., нервно-мышечное соединение, С. аксомышечный, С. нейромускулярный) — С. между аксоном мотонейрона и поперечно-полосатым мышечным волокном.
1) С., в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала;
2) аксо-аксональный С., обеспечивающий пресинаптическое торможение.
Синапс холинергический (s. cholinergica) — С., в котором медиатором является ацетилхолин.
Нервная система, как известно, состоит из нейронов. Эти особые клетки умеют принимать, хранить и обрабатывать информацию, они отвечают за связь организма с внешним миром и за работу всех систем этого организма. Память, внимание, мышление, воображение, творчество – всё это результаты работы нейронов. Однако вся эта многообразная деятельность не могла бы осуществиться, не будь у нейрона такого важного элемента, как синапс. В определённом смысле именно синапсы, а не сами нейроны, являются основой нервной системы.

Что такое синапс
Если сказать слишком упрощённо, то синапс – это место стыковки двух нервных клеток. Казалось бы, что здесь особенного? Но на самом деле синапс – это довольно сложное устройство, благодаря которому весь механизм сбора и обработки информации может исправно работать. Синапс – это то, что позволяет превратить простейшие сигналы и безусловные рефлексы в сложнейшие образцы мыслительной деятельности: представления, идеи, образы, произведения искусства, научные теории. Каково же строение синапса?
Строение синапса
Каждая нервная клетка имеет большое количество отростков. Все эти отростки, кроме одного, являются дендритами; это короткие и сильно разветвлённые образования, которые предназначены для приёма информации от других нейронов. Оставшийся длинный отросток называется аксоном; он отвечает за передачу информации от данной нервной клетки к следующей.
Соединяясь между собой отростками, нервные клетки образуют сложную сеть, по которой в разные стороны перемещаются сигналы. Разрозненные сигналы от периферической нервной системы попадают в центральную, где из них организм формирует целостную картину мира, решает, как ему поступать в дальнейшем, и посылает сигналы к нужным органам. Аксон нервной клетки может достигать внушительной длины – до полутора метров. И это только в организме человека. У жирафов аксоны в спинном мозге могут достигать и пяти метров. По-видимому, у более крупных вымерших животных, например, динозавров, аксоны нервных клеток в спинном мозге были ещё длиннее. Выходит, что нервные клетки являются самыми крупными клетками в организме.
Однако чаще всего напрямую от одной нервной клетки к другой сигнал пройти не может, потому что пространство между дендритами и аксоном заполнено межклеточным веществом. Чтобы нервная информация прошла от одного отростка к другому, нужно соорудить своеобразный мост. Такие мосты называются нейротрансмиттерами, или нейромедиаторами; образуются они в результате биохимических реакций и представляют собой белковые молекулы.

Сами нервные клетки очень маленькие – крупнейшие из них обычно не превышают длины 100 микрометров. Отростки нейронов, следовательно, имеют и вовсе микроскопические размеры. Однако даже на таком микроскопическом уровне строение синапса довольно сложное. Он состоит из трёх отделов. Первый – утолщение на конце аксона, называемое пресинапсической мембраной и необходимое для формирования нейромедиаторов. Второй отдел – аналогичное утолщение на конце дендрита, которое служит для приёма сигналов от нейромедиатора. Между ними находится третий отдел – сама синаптическая щель, в которой нейромедиаторы образуются.
Но строение синапса этим не ограничивается. На утолщении аксона имеются особые образования – синаптические пузырьки, которые содержат либо нейромедиатор, либо фермент, разрушающий нейромедиатор. А на утолщении дендрита имеются рецепторы, принимающие сигналы от конкретного нейромедиатора.
Данное строение синапса характерно для химического типа. Есть ещё электрические синапсы, имеющие несколько другую структуру. Нейромедиаторов они не образуют, поскольку электрические сигналы беспрепятственно проходят сквозь межклеточное вещество. При этом расстояние между мембранами в электрическом синапсе гораздо меньше, чем в химическом, благодаря чему давление межклеточного вещества более слабое. Кроме того, мембраны соединены так называемыми коннексонами – особыми белковыми образованиями.
Бывают ещё и смешанные синапсы, в которых химическая связь является фактором, усиливающим электрическую передачу сигнала.
Наиболее распространёнными являются химические синапсы, которые являются типовой разновидностью. Особенно велика их роль в нервной системе млекопитающих.

Аксоны и дендриты образуются в нервных клетках не сразу. Причём первым пробивается именно аксон, который начинает усиленно расти и прокладывать себе путь в окружающем пространстве. Так начинается рост самой нервной клетки. В конце концов аксон встречается с дендритами других нервных клеток и вместе с ними образует синапс.
Известно, что в нервной системе содержатся нейроны, не имеющие аксонов. Как работают такие клетки и для чего они нужны – пока никто не знает.
Интересно, что исследования работы нервных клеток учёные проводили на кальмарах. Их нервные клетки настолько большие, что видны невооружённым глазом. Это позволило вставлять в них электроды и измерять электрический потенциал в разных частях клетки. Исследователи Ходжкин, Элкс и Хаксли за такую работу в 1963 году удостоились Нобелевской премии.
Классификация синапсов
Существует несколько классификаций соединений нервных клеток. Первую из них мы рассмотрели выше – это деление на химические, электрические и смешанные синапсы. Также синапсы можно разделить по характеру передаваемого сигала: возбуждающие и тормозящие. Синапсы могут быть разделены и по месту расположения: центральные, находящиеся в головном мозге, и периферические, расположенные в периферической нервной системе.
Также синапсы делят в зависимости от производимых нейромедиаторов. Одни производят норадреналин, другие – ацетилхолин, серотонин, глутамат и другие. Всего существует около шестидесяти видов нейромедиааторов, каждый из которых несёт специфическую функцию. Так, норадреналин является возбуждающим веществом, он активизирует все системы организма, порождает чувство ярости. Дофамин – гормон счастья, который сообщает организму состояние блаженства, порождает позитивные эмоции; также он отвечает за познавательные процессы. Как переизбыток, так и недостаток нейромедиаторов приводит к различным нарушениям в нервной системе и организме в целом. Так, недостаток дофамина порождает депрессию, упадок сил, приводит к слабоумию. Переизбыток глутамата может привести к гибели нервных клеток.

Нейроны – это нервные клетки нашего организма, которые распространяют информацию по всему телу при помощи электрических сигналов. Это помогает управлять всеми жизненно важными функциями организма. Всего у человека имеется около 100 млрд нейронов.
Нейроны являются основой нервной системы. Она включает в себя головной и спинной мозг, нервы и нервные окончания.
Как устроены нейроны?
Все нейроны имеют особое строение. У каждого нейрона есть тело (второе название - сома) и похожие на щупальца длинный и короткие отростки. Короткие отростки называют дендритами. Они принимают информацию от других нейронов. Длинный отросток всего один и называется он аксоном. Главная его роль – передать информацию от своего нейрона другим нервным клеткам.
Аксоны, объединяясь друг с другом, образуют нервы. Каждый аксон покрыт миелиновой оболочкой - жироподобным белым веществом. Эта обертка позволяет электрическому сигналу быстрее и надежнее передвигаться по всей нервной системе.
Что такое синапс?
Все нейроны соединяются друг с другом по одной схеме: аксон одного нейрона соединяется с телом или дендритами второго нейрона.
Место сближения двух нейронов называется синапсом. На теле только одного нейрона насчитывается от 1200 до 1800 синапсов.
Важно отметить, что два нейрона на месте синапса не соприкасаются, между ними находится синаптическая щель. Через эту щель проходит специальное вещество – медиатор, которое и обеспечивает передвижение нервного импульса от одного нейрона ко второму.
Какие функции выполняют нейроны?
Несмотря на то, что у всех нейронов есть основные функции приема и передачи информации, ученые разделяют их на разные группы.
По функциям нейроны бывают:
1) Чувствительные или афферентные – это нейроны, которые самые первые получают информацию от ощущений, таких как вкус, запах, боль, звук, температура.
2) Вставочные или промежуточные нейроны оправдывают свое название тем, что находятся и проводят нервный сигнал между чувствительными и двигательными нейронами.
3) Двигательные или эфферентные – нейроны, которые проводят сигнал от мозга к мышцам или железам.
По количеству отростков нейроны делят на:
1) Униполярные – имеют один отросток. Он играет роль как дендрита, так и аксона.
2) Биполярные – обладают двумя отростками: один дендрит и один аксон. Эти нейроны участвуют в зрительной, слуховой и обонятельных системах.
3) Псевдоуниполярные – нейроны, от тела которых отходит один отросток, который позже раздваивается.
4) Мультиполярные нейроны обладают большим количеством дендритов и всего одним аксоном. Находятся в головном и спинном мозге.
Восстанавливаются ли нервные клетки?
Долгое время считалось, что нервные клетки не восстанавливаются. Однако учёные из Каролинского института выяснили, что в день образуется до 700 нейронов. Новые нервные клетки формируются из стволовых клеток. Эти клетки отвечают за созревание мозга плода в утробе матери.
Не восстанавливаются нервные окончания, и поэтому человек теряет способность ходить при серьезных повреждениях спинного мозга.
Мы можем помочь сами себе в восстановлении нервных клеток. Для этого решайте сложные задачи, принимайте решения, находите выход из непростых ситуаций. Это стимулирует работу мозга и заставляет нервные клетки быстрее восстанавливаться.
Синаптогенез - процесс формирования синапсов между нейронами в нервной системе. Синаптогенез происходит на протяжении всей жизни здорового человека, а взрыв формирования синапсов, т. н. избыточный синаптогенез (exuberant synaptogenesis), наблюдается на ранних стадиях развития головного мозга. [1] Синаптогенез особенно важен в ходе критического периода развития особи (в биологии развития, такого периода, когда нервная система особенно чувствительна к экзогенным стимулам), когда имеет место интенсивное протекание синаптического прунинга ввиду конкуренции нейронов и их синапсов за нейрональные факторы роста. Синапсы, которые не участвуют или участвуют слабо в обработке информации, как и неиспользуемые или малоиспользуемые отростки нейронов, не получат должного развития и будут отсеяны в дальнейшем. [2]
Содержание
- 1 Формирование нервно-мышечных синапсов
- 1.1 Функция
- 1.2 Происхождение и судьба клеток
- 1.3 Послесинаптическая дифференциация
- 1.3.1 Кластеризация
- 1.3.2 Синапс-специфическая транскрипция
- 1.3.3 Внесинаптическая репрессия
- 1.4 Досинаптическая дифференциация
- 1.5 Созревание синапса
- 1.5.1 Элиминация синапса
- 1.6 Специфика формирования синапсов
- 2 Формирование синапсов в ЦНС
- 2.1 Факторы, регулирующие синаптогенез в ЦНС
- 2.1.1 Сигналинг
- 2.1.2 Морфология
- 2.1 Факторы, регулирующие синаптогенез в ЦНС
- 3 Экологическое обогащение
- 4 Роль белкового семейства Wnt
- 4.1 Центральная нервная система
- 4.2 Нервно-мышечный синапс
- 5 Ссылки
Формирование нервно-мышечных синапсов [ | ]
Нервно-мышечное соединение (НМС) из всех типов синапсов наиболее хорошо изучено, оно представляет собой простую и доступную структуру для манипулирования и наблюдения. Этот синапс сам по себе состоит из трех клеток: мотонейрона, миоцита, и шванновской клетки (нейролеммоцита). В нормально функционирующем синапсе сигнал от мотонейрона вызывает деполяризацию мембраны миоцита, что происходит с помощью нейротрансмиттера ацетилхолина. Ацетилхолин высвобождается аксоном мотонейрона в синаптическую щель, где он достигает холинэргических рецепторов на сарколемме, мембране миоцита. Связывание ацетилхолина с рецепторами приводит к открытию ионных каналов, деполяризации мембраны и последующему каскаду химических реакций, результатом чего будет сокращение мышцы. Снаружи синапс инкапсулирует и изолирует миелиновая оболочка шванновской клетки. [3] Еще одним важным элементом центральной и периферической нервной системы являются астроциты. Хотя первоначально и полагалось, что их функция лишь структурная, сейчас известно, что они играют важную роль в функциональной пластичности синапсов. [4]
В ходе развития каждый из трех типов вышеупомянутых клеток возникает из разных регионов развивающегося эмбриона. Отдельные миобласты развиваются из мезодермы и объединяются, формируя полиэнергидное (многоядерное) мышечное волокно. Во время или вскоре после образования волокна, мотонейроны нервной трубки начинают формировать первые контакты с ним. Шванновские клетки возникают из нервного гребня и направляются вместе с аксонами к месту назначения. По достижении мышечного волокна, они образуют свободное, немиелинизированное покрытие иннервирующих мышцы аксонов. Движение аксонов (а т. о. и клеток Шванна) управляется конусом роста, нитевидной проекцией аксона, которая активно ориентируется на нейротрофины, высвобождаемые мышечным волокном.
Исследование паттернов образования и распределения нервно-мышечных синапов показывает, что существенная часть соединений локализуется в средней точке мышечного волокна. Хотя может показаться, что аксоны изначально нацеливаются на этот локус, несколько факторов говорят об обратном. По-видимому, после первоначального контакта с аксоном, молодое волокно продолжает расти симметрично от точки иннервации. В сочетании с тем фактом, что высокая плотность холинэргических рецепторов является результатом контакта с аксоном, а не его причиной, паттерны формирования мышечных волокон можно связать как с индивидуальным миопатическим ростом, так и с иннервацией волокна аксонами.
Трансдукция сигнала по первичному контакту между мотонейроном и мышечным волокном начинается почти сразу, но передаваемый импульс очень слабый. Существуют свидетельства того, что Шванновские клетки, с помощью небольших сигнальных молекул, могут усиливать импульсы путем увеличения интенсивности спонтанного высвобождения нейромедиаторов. [5] Примерно через неделю мотонейрон и мышечное волокно проходят процесс дифференциации, чем завершается формирование полностью функционального синапса. Иннервация изначальным аксоном имеет решающие значение, поскольку последующие за ним аксоны имеют высокую склонность к формированию контактов с уже устоявшимся, стабилизированным синапсом.
Наиболее заметное изменение в мышечном волокне, возникающее после контакта с мотонейроном, - это повышенная концентрация холинэргических рецепторов на плазмалемме в месте контакта нейрона и миоцита. Это увеличение числа рецепторов обеспечивает более эффективную передачу синаптических сигналов, что в свою очередь ведет к формированию более развитого синапса. Плотность рецепторов составляет >10,000/мкм 2 в центре и приблизительно 10/мкм 2 на периферии. Такая высокая концентрация холинэргических рецепторов в синапсе достигается посредством их кластеризации, повышенной регуляции экспрессии соответствующих генов в ядрах пост-синаптических клеток и пониженной регуляции экспрессии этих генов в ядрах пре-синаптических клеток. Сигналами, которые инициируют послесинаптическую дифференциацию, могут быть нейротрансмиттеры, высвобождаемые непосредственно из аксона на мышечное волокно, или они могут возникать сторонним образом, попадая в синаптическую щель извне. [6]
Кластеризация на пост-синаптической мембране происходит во многом благодаря протеогликану Агрину (Agrin), который экскретируется аксоном мотонейрона и способствует мультимеризации рецепторов. Агрин связывается с рецепторной мышечно-специфичной киназой (MuSK) на пост-синаптической мембране, а это в свою очередь приводит к последующей активации цитоплазматического белка Рапсина (Rapsyn). Именно рапсин содержит домены, позволяющие ему кластеризовать на мембране множество холинэргических рецепторов: мышечные клетки мутантных на этот белок мышей не были способны формировать кластеры холинэргических рецепторов.
Повышенная концентрация холинэргических рецепторов достигается не только за счет манипуляций с уже существующими синаптическими компонентами. Аксон путём сигналинга способен на уровне процесса транскрипции осуществлять регуляцию экспрессии генов в ядре мышечной клетки. Всё это также происходит для того, чтобы повысить интенсивность локальной кластеризации рецепторов на мембране. В частности, в сигналинге задействованы две молекулы: белок семейства кальцитонинов (Calcitonin gene-related peptide) и представитель семейства белков-нейрегулинов (названных так за свою роль в регуляции нейрональных молекулярных процессов). Эти два белка вызывают каскад химических реакций с участием различных киназ, что в конечном итоге ведет к активации транскрипции генов холинэргических рецепторов. [7]
Новообразованные синапсы способны генерировать электрические сигналы, таким образом воздействующие на окружающие внесинаптические клетки, что те существенно сокращают экспрессию генов холинэргических рецепторов. Точность импульса, посылаемого аксоном на пост-синаптическую мембрану, достигается именно за счет одновременного снижения концентрации рецепторов вне синапса, и повышения их количества внутри него. Поскольку синапс начинает получать входную информацию почти сразу после образования нервно-мышечного соединения, аксон быстро генерирует потенциал действия и высвобождает ацетилхолин. Деполяризация, вызванная связыванием ацетилхолина с его рецепторами, инициирует сокращение мышцы, и, одновременно с этим, ингибирование транскрипции рецепторных генов в клетках мышечного волокна вокруг синапса. Следует заметить, что эффект действует на расстоянии, а рецепторы, сидящие непосредственно на пост-синаптической мембране, репрессии не подвергаются.
Хотя механизмы, регулирующие досинаптическую дифференциацию, неизвестны, изменения, происходящие в развивающейся аксональной терминали, охарактеризованы достаточно хорошо. Увеличивается объем и площадь синаптического контакта, объем везикул, концентрация везикул в активной зоне синапса, а также поляризация пресинаптической мембраны. Эти изменения, как считается, опосредованы нейротрофином и различными молекулами клеточной адгезии, высвобождающимися миоцитом; это подчеркивает важность связи между мотонейроном и мышечным волокном в процессе синаптогенеза. Считается, что именно изменение экспрессии генов и перераспределение составляющих компонентов синапса - ключевые моменты до- и послесинаптической дифференциации. В пользу этого также свидетельствует усиление экспрессии генов везикулярных белков вскоре после формирования синапса, а также их аклокализация в аксональной терминали.
Незрелые синапсы обнаруживают множественную иннервацию ввиду высокой склонности новых аксонов связываться с уже существующими синапсами. По мере созревания, синапсы обособляются и, в конечном счете, на входе остается только один аксон; это происходит в процессе, называемом элиминация синапса. Кроме того, путём инвагинации постсинаптическая сторона увеличивает площадь принимающей нейротрансмиттеры поверхности. На ранних этапах онтогенеза, шванновские клетки формируют рыхлую, немиелинизирующую оболочку, покрывающую группу синапсов, однако по мере роста и развития, пределы компетенции отдельно взятого нейролеммоцита сужаются до одного синапса.
Элиминация синапса, как один из механизмов синаптического прунинга, предполагает под собой процесс, напрямую зависящий от активности конкурирующих между собой аксонов. В случае конкретно взятого синапса, если частота импульсов на входе достаточна для создания потенциала действия, то это будет провоцировать экспрессию генов белков-синаптотрофинов в ядре пост-синаптического миоцита, и эти белки будут поддерживать данный синапс. Возникает механизм положительной обратной связи, недоступный менее востребованным синапсам, вследствие этого обреченным на элиминацию. Также было высказано предположение, что в дополнение к синаптотропинам, высвобождаемым в активных синапсах, деполяризация их постсинаптической мембраны вызывает высвобождение синаптотоксинов, ингибирующих аксоны в неактивных синапсах.
Формирование синапсов в ЦНС [ | ]
Данные, полученные в исследованиях нервно-мышечных синапсов, можно попытаться экстраполировать на процессы синаптогенеза в ЦНС, даже несмотря на то, что исследования последних имеют намного более поздний характер. Эти два типа соединений имеют много общего. Оба типа синапса имеют консервативное базовое строение: пре-синаптическая нервная терминаль, синаптическая щель и принимающая постсинаптическая клетка. Оба типа синапса содержат высокую концентрацию везикул в активном центре, кластеры рецепторов на принимающей мембране, и, наконец, оба они изолированы специальными глиальными клетками. Процессы, происходящие в них, также достаточно консервативны. Это и кластеризация рецепторов, и локализованное повышение интенсивности синтеза белка в активных центрах, и синаптический прунинг путем элиминации малоактивных синапсов.
Несмотря на вышеобозначенные сходства, существует принципиальная разница между этими двумя соединениями. Поскольку постсинаптической клеткой уже не выступает миоцит, в корне отличаются действующие рецепторы и их нейротрансмиттеры. Что еще более важно, нейроны в пределах ЦНС иннервированы многими другими нейронами одновременно, и должны уметь интегрировать и обрабатывать большое количество информации на входе, когда как мышечные волокна иннервируются одним нейроном и сокращаются в ответ на одиночный импульс. В сочетании с пластичностью, характерной для нейронных связей, нейронная сеть в ЦНС имеет огромный потенциал для усложнения.
Гомологом ацетилхолина и его рецепторов в ЦНС является глутамат и N-метил-D-аспартат-рецепторы (NMDA). Было показано, что активация NMDA-рецепторов инициирует каскад химических реакций, ведущих к формированию и стабилизации синапса. Повышенный уровень активности NMDA-рецепторов во время развития синапса позволяет увеличить приток кальция, который выступает в качестве вторичного сигнала. Помимо этого, факторами транскрипции активируются гены первичного ответа (ГПО), играющие важную роль в таких процессах, как, например, долговременная потенциация. [8] Функция рецепторов NMDA также связана с эстрогеновыми рецепторами в гиппокампе. Эксперименты, проведенные с эстрадиолом, показывают, что воздействие эстрогена значительно увеличивает плотность синапсов и концентрацию белка. [9]
Передача нервного импульса во время синаптогенеза зависит не только от активности отдельно взятого синапса, но и от окружающей этот синапс среды. Например, нейротрофический фактор головного мозга (BDNF) - продуцируется головным мозгом и регулирует несколько функций в развивающемся синапсе, включая усиление высвобождения нейротрансмиттеров, повышение концентрации везикул и биосинтез холестерина. Холестерин является важной структурной составляющей липидного бислоя, выступающим как место действия самых разных сигнальных взаимодействий. Мутантные на наличие BDNF организмы обнаруживают значительные дефекты в росте нейронов и формировании синапсов. [10] Помимо нейротрофинов, важную роль в процессе синаптогенеза играют также молекулы клеточной адгезии. Во многом синаптогенез зависит от корректного локального связывания одной адгезионной молекулы с другой. Показано, что дефекты в генах, ирующих белки-нейролигины, одни из важнейших молекул клеточной адгезии, играющие роль в процессе стабилизации синапса, связаны с некоторыми случаями аутизма и умственной отсталости. [11] Наконец, многие из этих сигнальных процессов могут регулироваться с помощью матриксных металлопротеиназ (ММП).
Высоинамичными сайтами, принимающими одновременно несколько входящих импульсов, являются дендритные шипики. Этот морфологический динамизм обусловлен специфической регуляцией актинового цитоскелета, что, в свою очередь, позволяет регулировать образование синапсов. [12] Морфологически, дендритные шипики можно разделить на три основных вида: филоподии, тонкие шипики и грибовидные шипики. Филоподии инициируют контакт с аксонами других нейронов. Филоподии новообразованных нейронов имеют тенденцию к образованию контактов с аксонами, иннервирующими множество синапсов, когда как филоподии более старых нейронов связываются преимущественно с аксонами, иннервирующими одиночный синапс. Благодаря высокой динамичности, одни типы шипиков способны переходить в другие, например, филоподии легко трансформируются в грибоподобные шипики, которые являются первичными сайтами глутаматных рецепторов. [13]
Экологическое обогащение [ | ]
Экологическое обогащение - термин, обозначающий стимуляцию развивающегося мозга физическим и социальным окружением. Экологическое обогащение приводит к тому, что эффективность процессов синаптогенеза существенно увеличивается. Так, выращенные в среде с таким эффектом крысы показывают увеличение количества синапсов на 25% в сравнении с контролем. [14] [15] Этот эффект наблюдается независимо от того, возникает ли стимулирующая среда сразу после рождения, [16] вскоре после отлучения, [17] или во время взросления. [18] Не только для пирамидальных, но и для звездчатых нейронов экологическое обогащение вызывает всплеск синаптогенеза. [19]
Роль белкового семейства Wnt [ | ]
В белковом семействе Wnt существует несколько эмбриональных морфогенов (веществ, определяющих морфогенез тканей), которые способствуют раннему формированию структур развивающегося эмбриона. Недавно появившиеся данные показывают вклад морфогенов этого семейства в синаптогенез и пластичность, причем семейство вносит вклад в синаптогенез как синапсов ЦНС, так и нервно-мышечных синапсов.
В синапсах мозжечка Wnt-семейство индуцирует созревание пре- и постсинаптических окончаний. Эта область мозга содержит три основных типа нейронов: клетки Пуркинье, гранулярные клетки и клетки мшистых волокон. У клеток Пуркинье Wnt-3 стимулирует рост как дендритов, так и аксонов. [20] [21] Гранулярные клетки экспрессируют Wnt-7а, который индуцирует рост и ветвление аксонов в клетках мшистых волокон. Ретроградная секреция Wnt-7a, направленная на мшистые волокна,вызывает увеличение конуса роста аксонов путем увеличения числа микротрубочек. Кроме того, ретроградный Wnt-7a-сигналинг вызывает накопление синаптических пузырьков и пресинаптических белков в активной зоне синапса. Внт-5А выполняет аналогичную функцию у постсинаптических гранулярных клеток; Wnt-5-сигналинг стимулирует сборку рецепторов на мембране, а также кластеризацию на мембране каркасного белка PSD-95.
В гиппокампе белки семейства Wnt, в сочетании с электрической активностью клеток, также способствует формированию синапсов. Wnt7b экспрессируется в созревающих дендритах, помимо этого, в формирующихся синапсах возрастает экспрессия рецепторов семейства Frizzled (Fz), специфических для Wnt-белков. Глутаматные NMDA-рецепторы активируются в ответ на экспрессию белков Wnt-2-типа. Долговременная потенциация вследствие активации NMDA-рецепторов и последующая Wnt-экспрессия ведет к кластеризации рецепторов Fz-5 типа на мембране пост-синаптических клеток. Кроме того, Wnt-7a и Wnt-2 типы сигналинга после опосредованной таким образом долговременной потенциации увеличивают интенсивность процесса ветвления дендритов, а также регулируют синаптическую пластичность. Блокирование экспрессии белков Wnt-семейства закономерно приводит к ингибированию этих процессов. [22]
Подобные механизмы, опосредованные Wnt-семейством в ЦНС, хорошо прослеживаются и в нервно-мышечных соединениях. У Дрозофил мутации Wnt-5-рецептора в нервно-мышечных синапсах приводят к снижению количества и плотности активных центров. Основным нейротрансмиттером в этой системе является глутамат. Белки Wnt-семейства необходимы для правильной локализации глутаматергических рецепторов на постсинаптической мембране миоцитов. Мутации в белках этого семейства приводят к снижению интенсивности волны деполяризации после передачи импульса с аксона на мышечное волокно.
У позвоночных в нервно-мышечных синапсах экспрессия белка Wnt-11r способствует кластеризации холинэргических рецепторов на постсинаптической мембране мышечных клеток. Белок Wnt-3 экспрессируется мышечными волокнами и секретируется ретроградно на моторные нейроны. В моторных нейронах, Wnt-3 работает в паре с Агрином для стимуляции расширения конуса роста, ветвления аксональных терминалей и кластеризации синаптических пузырьков.
Читайте также:
