
Состояние двигательных нервных центров
Одностороннее проведение возбуждения. В нервных центрах прохождение импульсов возбуждения по цепи нейронов осуществляется только в одном направлении: от чувствительного нерва через промежуточные к двигательному, а от двигательного — к органу-исполнителю. Это обусловлено односторонней синаптической передачей импульсов от одной нервной клетки к другой с помощью медиатора, который выделяется концевым аппаратом аксона и содержится лишь в пресинаптической щели. В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление. Это свойство обеспечивает координирующую роль центральной нервной системы и способствует замыкательной функции условно-рефлекторных связей.
Замедление проведения возбуждения.
Это свойство называется центральной задержкой, или латентным (скрытым) периодом рефлекса. Центральная задержка обусловлена более медленным проведением нервных импульсов через синапсы.
Интервал от начала раздражения рецептора до появления ответной реакции — 0,2–0,5 секунды. Чем сложнее рефлекс, тем длительнее центральная задержка. Дрессировщику необходимо учитывать скрытый период при выработке условного рефлекса и подкрепляющий раздражитель надо применять не раньше 0,5 секунды после воздействия сигнального раздражителя. Скрытый период рефлекса увеличивается при утомлении и заболевании животного.
Присуще всем нервным центрам и характеризуется тем, что на короткое раздражение проявляется длительная ответная реакция, т. е. рефлекторный акт длится еще некоторое время после того, как раздражение рецептора прекращено. Такое последействие объясняется тем, что нервные импульсы от рецепторов поступают к двигательным центрам по различным нервным путям неодновременно: по коротким быстрее, чем по длинным. Запаздывающие импульсы поддерживают возбужденное состояние соответствующего нервного центра. Остаточное возбуждение в нервных центрах может сохраняться до 2 секунд и тем самым способствовать лучшей замыкательной функции при образовании условных рефлексов. Кроме того, в практике дрессировки нужно помнить, что после окончания действия любого раздражителя требуется некоторое время для спада остаточного возбуждения нервного центра и освобождения рефлекторных путей для новой рефлекторной деятельности.
Происходит путем накопления слабых допороговых раздражений до критического потенциала, способного вызвать возбуждение нервного центра. Различают пространственную и временную суммацию. Пространственная суммация наблюдается в случае одновременного воздействия раздражителей допороговой силы на несколько чувствительных нейронов. Поступившие импульсы слабой силы суммируются в нервном центре и вызывают возбуждение. Суммация во времени происходит при активизации одного и того же чувствительного нейрона серией последовательных раздражений допороговой силы. Слабые импульсы от предыдущих раздражений накладываются друг на друга, суммируются и вызывают рефлекс. Суммация в пространстве и во времени в нервных центрах происходит одновременно. Они дополняют и усиливают друг друга и, например, увеличивают обонятельную и слуховую чувствительность у собаки, что так необходимо при работе по чутью, когда исключительно слабые раздражения запаховых частиц (1 молекула в одном литре воздуха) вызывают запаховые ощущения и соответствующие ответные реакции.
Свойство нервных центров изменять частоту и силу передающихся импульсов, т. е. трансформировать. Проявляется в активизации и перестройке нейронов на более высокий или более низкий ритм и изменении их лабильности, обеспечивает взаимодействие и установление связи между различными нервными центрами и другими отделами нервной системы, что имеет важное значение в замыкании условно-рефлекторных связей при дрессировке собак.
Свойство нервных центров обеспечивать высокую возбудимость и эффективность рефлекторной деятельности нервной системы. Сущность облегчения заключается в том, что после каждого раздражения в нервном центре повышается возбудимость к повторным раздражениям, следующим через небольшие промежутки времени. Один поток импульсов как бы облегчает действие другого и способствует образованию условного рефлекса.
Способность одних нервных центров повышать возбудимость других и вступать во взаимодействие через обмен и передачу импульсов возбуждения между центрами по нейронным путям. Процесс образования временной связи рассматривается как результат взаимодействия двух очагов возбуждения в коре, следствием чего является проторение пути между этими корковыми пунктами. Проходимость пути обусловлена функциональными и морфологическими изменениями в синапсах.
При сильном и длительном раздражении импульсы, поступившие в нервную систему, не ограничиваются возбуждением одного рефлекторного центра, а распространяются на другие центры. Чем сильнее раздражение, тем больше нервных центров охватывает иррадиация. Возбуждение большого количества различных нервных центров позволяет отобрать из них наиболее нужные и установить между ними новые функциональные связи — условные рефлексы. Большинство двигательных условных рефлексов формируется благодаря иррадиации возбуждения. Излишняя иррадиация возбуждения центральной нервной системы нарушает уравновешенность поведенческих реакций, приводит к расстройству рефлекторной деятельности. Иррадиация возбуждения ограничивается и уравновешивается торможением.
Тормозной процесс — необходимое условие в координации нервной деятельности. Торможение возникает в определенных нервных структурах под влиянием волны возбуждения, которая подавляет другое возбуждение. При торможении выключается деятельность ненужных в данный момент органов и предохраняется от чрезмерного перенапряжения работа нервных центров. В нервной системе вырабатываются тормозные условные рефлексы: прекращения нежелательных действий, выдержки, дифференцировки и др.
Преобладание активности одних нервных центров над активностью других.
Доминирующий (господствующий) очаг возникает при определенном функциональном состоянии нервных центров. Одно из условий его образования — повышенная возбудимость нервных клеток одного или нескольких нервных центров под влиянием определенных нервных и гуморальных факторов. Возбудимость доминирующего очага усиливается суммированием импульсов, поступающих из других нервных центров, рефлекторная деятельность которых подавляется, а рефлекторная деятельность доминирующего очага заметно усиливается. Установившееся доминирование при инстинктах может быть продолжительным состоянием, которое определяет поведение животного на тот или иной срок.
Доминанту можно наблюдать и в условно-рефлекторной деятельности. Доминирующий очаг возбуждения в коре головного мозга притягивает к себе импульсы нервных центров других очагов возбуждения и тем самым способствует суммации, облегчению, проторению и замыканию условно-рефлекторных связей на сигналы дрессировщика. Доминанта по теории академика И.П. Павлова имеет главенствующее значение в механизме образования и проявления условных рефлексов. У собаки всегда сильно проявляются стойко выработанные навыки, доминирующие над другими рефлексами и обеспечивающие безотказное управление собакой при отвлекающих раздражителям. Дрессировщику необходимо учитывать свойство доминанты при подготовке собак.
Схождение поступающих по чувствительным путям импульсов возбуждения в одном промежуточном или двигательном центре. В центральной нервной системе в 4–5 раз больше чувствительных путей, чем двигательных. Поэтому к одному и тому же двигательному центру могут подходить импульсы возбуждения по многим путям. Эта особенность прохождения возбуждения по нервным центрам противоположна иррадиации и является основой для концентрации возбуждения в отдельных пунктах коры головного мозга. Конвергенция обеспечивает специализацию условного рефлекса и формирование навыка на сложные и комплексные раздражители.
Проявляется при сочетании особенно сильных раздражителей, дающих эффект меньше суммы величин этих реакций на каждый раздражитель в отдельности. Это свойство противоположно суммации. Окклюзия и пространственная суммация постоянно взаимодействуют между собой. При слабом возбуждении проявляется суммация, при сильных раздражениях — окклюзия. В дрессировке собак ошибочно применение сильных раздражителей для ускорения первоначального условного рефлекса. На громкие команды и сильные подкрепления вырабатываются, как правило, слабые условные рефлексы.
Способность перестройки функций нервных центров. Сформировавшиеся в процессе эволюции рефлекторные акты, реакции поведения и инстинкты могут перестраиваться, а нервные центры изменять свои функции. Опытами установлено, что перестройка функций нервных центров у животных происходит под регулирующим влиянием коры полушарий головного мозга. Благодаря этому свойству при изменении условий жизни меняется поведение животных, их привычки и навыки: собака и кошка могут мирно уживаться, инстинкт преследования собакой диких животных по следам заменяется реакцией поиска человека по его запаховому следу, вредные привычки, нежелательные связи у собаки могут быть исправлены систематической дрессировкой. Пластичность нервных центров позволяет при дрессировке вырабатывать сложные навыки в виде динамических стереотипов.
Нервные центры обладают свойством переходить в состояние возбуждения только при относительно длительном раздражении. Возбудившись, они сохраняют это состояние возбуждения в течение определенного времени. Такое явление И. П. Павлов назвал инертностью. Способность нервных центров длительно сохранять в себе следы возбуждения и торможения, сильно выражена в нервных клетках коры полушарий головного мозга. И. П. Павлов говорил, что если бы у нервных клеток не было инертности, у нас не было бы никакой памяти, никакой выучки, не существовало бы никаких привычек. У животных существует два вида памяти: кратковременная и долговременная. Оба вида памяти обеспечивают возможность выработки у животных условных рефлексов и формирования стойких навыков. Кратковременная память у собаки проявляется в течение нескольких минут, долговременная, или долгосрочная, память — через много дней, месяцев и даже лет. Оба вида памяти у животных выражаются в репродукции образа воспринимаемых отдельных раздражителей или целостного объекта.
Нервные центры обладают свойством постоянно находиться в состоянии незначительного возбуждения при относительном рефлекторном покое. Они постоянно посылают импульсы, обеспечивающие тоническое сокращение скелетной мускулатуры. Тонус нервных центров поддерживается действием гуморальных веществ и непрерывным потоком импульсов, поступающих от рецепторов. Огромное значение в поддержании мышечного тонуса имеют нервные центры продолговатого, среднего и промежуточного мозга. Тоническое состояние нервных центров и мышечной системы обеспечивает устойчивую выработку условных рефлексов и хорошую работоспособность собаки. Состояние рабочего тонуса поддерживается правильно организованной дрессировкой, систематической тренировкой и регулярным использованием собаки на службе.
Перечисленные свойства нервных центров обеспечивают их функциональное назначение в нервной системе и организме.
из книги Арасланов Филимон, Алексеев Алексей,
Шигорин Валерий "Дрессировка собак"
при использовании материалов
ссылка на ресурс wolcha.ru
Новость отредактировал: maugli - 11-10-2019, 07:19
Нервный центр – это совокупность нейронов, обеспечивающих регуляцию какого-либо конкретного физиологического процесса или функции.
Нервный центр в узком смысле – это совокупность нейронов, без которых данная конкретная функция не может регулироваться. Например, без нейронов дыхательного центра продолговатого мозга дыхание прекращается. Нервный центр в широком смысле — это совокупность нейронов, которые участвуют в регуляции конкретной физиологической функции, но не являются строго обязательными для ее осуществления! Например, в регуляции дыхания кроме нейронов продолговатого мозга участвуют нейроны пневмотаксического центра варолиевого моста, отдельные ядра гипоталамуса, кора больших полушарий и другие образования головного мозга.
Все нейроны нервного центра разделяют на 2 неравные по количеству и качеству группы.

Первая группа – нейроны центральной зоны. Это наиболее возбудимые нейроны, которые возбуждаются в ответ на поступление порогового (для нервного центра) сигнала. Таких нейронов около 15-20%, и они не обязательно располагаются в середине нервного центра, как это изображено на рис.1. Особенностью их является то, что они имеют на своем теле больше синаптических терминалей от сенсорных и вставочных нейронов.
Вторая группа – нейроны подпороговой каймы. Это менее возбудимые нейроны, которые не возбуждаются в ответ на поступление пороговых им-пульсов, но при действии более сильных раздражителей они возбуждаются и включаются в работу нервного центра, обеспечивая ее усиление. Таких нейронов большинство (80-85%), и они не обязательно располагаются на периферии нервного центра, но все имеют значительно меньше синаптических терминалей от сенсорных и вставочных нейронов по сравнению с нейронами центральной зоны.
На рис. 1 нейроны центральной зоны условно поставлены в центр внутреннего круга (А), а нейроны подпороговой каймы – в пространство между внутренним и наружным кругами (Б). Таким образом, если к нервному центру по афферентному входу (В) придет пороговый импульс, то возбудятся три нейрона центральной зоны, а на десяти нейронах подпороговой каймы потенциалы действия не возникнут, но появится местная деполяризация – возбуждающий постсинаптический потенциал (ВПСП).
От структуры нервного центра зависят его свойства, а они, в свою очередь, влияют на процесс проведения возбуждения через нервный центр, на его скорость и степень выраженности. От свойств нервных центров во многом зависит процесс распространения возбуждения по ЦНС, что имеет важное значение в интегративной деятельности организма.
Свойства нервных центров обусловлены описанной выше нейронной организацией нервного центра, а также химическим способом передачи возбуждения в синапсах. При электрическом способе передачи возбуждения нервные центры не имели бы подобных свойств.
Свойства нервных центров: 1 одностороннее проведение возбуждения; 2 задержка проведения возбуждения; 3 суммация; 4 облегчение; 5 окклюзия; 6 мультипликация; 7 трансформация; 8 последействие; 9 посттетаническая потенциация; 10 утомление; 11 тонус; 12 высокая чувствительность к изменению состояния внутренней среды организма; 13 пластичность.
3) Суммация – это возникновение возбуждения в нервном центре при поступлении к нему нескольких допороговых импульсов, каждый из которых в отдельности не может возбуждения (рис. 2). Фактически этот процесс происходит на нейронах подпороговой каймы. Различают два вида суммации: пространственную и временною.

Пространственная суммация возникает в том случае, когда к нервному центру (к его нейронам) приходят одновременно, несколько допороговых импульсов. На рисунке 2А видно, что к нейрону подпороговой каймы, имеющему пороговый потенциал 30 мВ одновременно по пяти различным афферентным входам (их аксоны обозначены сплошной линией) приходят пять импульсов, каждый из которых деполяризует мембрану нейрона на 5 мВ (то есть возникают пять отдельных ВПСП). В этом случае возбуждение нейрона не наступает, так как суммарная деполяризация мембраны нейрона составляет лишь 25 мВ (суммированный ВПСП мал для достижения КУД). Но если к нейрону придет еще один подобный импульс по шестому входу (его аксон обозначен пунктирной линией), то суммированный ВПСП будет достаточен по величине и мембрана нейрона в зоне аксонного холмика деполяризуется до критического уровня, в результате чего нейрон из состояния покоя перейдет в состояние возбуждения. На постсинаптической мембране происходит суммация ВПСП в пространстве.
Временная (последовательная) суммация возникает в том случае, когда к нейронам нервного центра по одному афферентному входу приходит не один, а серия импульсов с очень небольшими по времени межимпульсными промежутками (рис. 2Б). Два механизма временной суммации:
1) интервалы между отдельными импульсами настолько малы, что за это время медиатор, выделившийся в синаптическую щель, не успевает полностью разрушиться и вернуться в пресинаптический аппарат. В этом случае возникает постепенное накопление медиатора до критического объема, необходимого для возникновения достаточного по амплитуде ВПСП, а значит, и для возникновения ПД;

2) интервалы между отдельными импульсами настолько малы, что возникший за это время на постсинаптической мембране ВПСП не успевает исчезнуть и усиливается за счет новой порции медиатора – суммируется. На постсинаптической мембране происходит суммация ВПСП во времени.
4) Облегчение – это увеличение количества возбужденных нейронов в нервном центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному, а по двум или более афферентным входам. На рис. 3 рассмотрен случай, когда при отдельном раздражении первого афферентного входа возбуждается только три нейрона центральной зоны (А), а на пяти нейронах подпороговой каймы (Б) возникают ВПСП. Если раздражать отдельно только второй афферентный вход, то возбуждены будут пять нейронов (Г), а четыре нейрона подпороговой каймы (Д) не возбудятся. Раздражая и первый, и второй афферентные входы одновременно (!), мы ожидаем вовлечения в процесс возбуждения восьми нейронов. И они, естественно, будут возбуждаться, но кроме них (сверх ожидания!) могут возбуждаться еще некоторые нейроны подпороговой каймы. Это произойдет потому, что один или несколько нейронов подпороговой каймы являются общими как для первого, так и для второго афферентных входов (в нашем случае это два нейрона, обозначенные буквой В), и при одновременном поступлении возбуждения к этим нейронам дни возбудятся за счет возникновения пространственной суммации.

5) Окклюзия – это уменьшение количества возбужденных нейронов в нервной центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному. а по двум или более афферентным входам (рис. 4).
6) Мультипликационное возбуждение (мультипликация) заключается в том, что по разветвлениям аксона вставочного нейрона возбуждение поступает одновременно не на один, а на несколько моторных нейронов (рис. 6). В связи с этим эффект на рабочем органе усиливается в несколько раз, или в работу вовлекаются не одна, а несколько рабочих структур, Это свойство особенно ярко проявляется в ганглиях автономной (вегетативной) нервной системы.

7) Трансформация ритма возбуждения – это изменение частоты импульсов на выходе из нервного центра по сравнению с частотой импульсов на входе в нервный центр.
а) наличием дублирующих цепей вставочных нейронов, связывающих сенсорные и моторные нейроны;
б) разным количеством синапсов в каждой из этих цепей.


Например, на рис.7 представлены два варианта трансформации, которые, на первый взгляд, не отличаются друг от друга, так как в том и в другом случае показаны две дополнительные цепи вставочных нейронов (кроме прямого пути), с помощью которых возбуждение может передаваться по цепи нейронов А—Б—В. Рассмотрим эти схемы.
Вариант 1. Верхняя цепь состоит из двух дополнительных вставочных нейронов, а значит, по сравнению с прямым путем передачи возбуждения с нейрона Б на нейрон В, имеет два дополнительных синапса. Поэтому возбуждение, проходя по верхней цепи, задержится на 2 мс (время синаптической задержки в одном синапсе составляет
1 мс) и придет на нейрон В после того, как пройдет возбуждение по прямому пути. В нижней цепи три дополнительных вставочных нейрона (то есть три дополнительных синапса), значит, возбуждение будет доходить до нейрона В еще дольше, чем по верхней цепи (задержка составит 3 мс). Следовательно, по нижней цепи возбуждение на нейрон В придет после того, как пройдет возбуждение по верхней цепи. В результате на один импульс, пришедший по сенсорному нейрону А, на моторном нейроне В возникнет три потенциала действия (трансформация 1:3).
Вариант 2. В этом случае и верхняя и нижняя цепи вставочных нейронов состоят из двух дополнительных нейронов. Возбуждение по обеим цепям придет к нейрону В одновременно в виде одного потенциала действие, который появится на нейроне В только после прохождения возбуждения к нему от нейрона Б по прямому пути. В этом варианте мы тоже получим трансформацию ритма, но уже в соотношении 1:2.
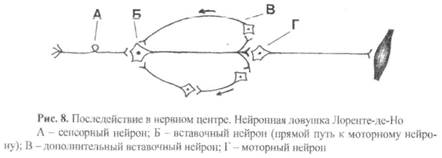
8) Последействие – это продолжение возбуждения моторного нейрона в течение некоторого времени после прекращения действия раздражителя.
Другими причинами последействия могут быть:
а) возникновение высокоамплитудного ВПСП, в результате которого возникает не один, а несколько потенциалов действия то есть ответ длится большее время;
б) длительная следовая деполяризация постсинаптической мембраны, в результате чего возникают несколько потенциалов действия, вместо одного.

9) Посттетаническая потенциация (синаптическое облегчение) – это улучшение проведения в синапсах после короткого раздражения афферентных путей.
Если в качестве контроля вызвать одиночное раздражение афферентного нерва тестирующим раздражителем (рис. 9А), то на моторном нейроне мы получим ВПСП вполне определенной амплитуды (в нашем случае 5 мВ). Если после этого тот же афферентный нерв раздражать некоторое время серией частых импульсов (рис. 9Б), а потом вновь подействовать тестирующим раздражителем (рис. 9В), то величина ВПСП будет больше (в нашем случае 10 мВ). Причем она будет тем больше, чем более частыми импульсами мы раздражали афферентный нерв.
Длительность синаптического облегчения зависит от свойств синапса и характера раздражения: после одиночных стимулов оно выражено слабо, после раздражающей серии потенциация (облегчение) может продолжаться от нескольких минут до нескольких часов. Объясняется он тем, что при частом раздражении афферентного волокна в его пресинаптической терминали (окончании) накапливаются ионы кальция, а значит, улучшается выделение медиатора. Кроме того, показано, что частое раздражение нерва приводит к усилению синтеза медиатора, мобилизации пузырьков медиатора, к усилению синтеза белков-рецепторов на постсинаптической мембране и увеличению их чувтствительности. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах.
10) Утомление нервного центра (посттетаническая депрессия, синаптическая депрессия) – это уменьшение или прекращение импульсной активности нервного центра в результате длительной стимуляции его афферентными импульсами (или произвольного вовлечения его в процесс возбуждения посредством импульсов, идущих из коры больших полушарий). Причинами утомления нервного центра могут быть:
• истощение запасов медиатора в афферентном или вставочном нейроне;
• снижение возбудимости постсинаптической мембраны (то есть мембраны моторного или вставочного нейрона) из-за накопления, например, продуктов метаболизма.
Утомляемость нервных центров продемонстрировал Н.Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения п. tibialis и п. peroneus. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги. Синаптическая депрессия при длительной активации центра выражается в снижении постсинаптических потенциалов.
11) Тонус нервного центра – это длительное, умеренное возбуждение нервного центра без видимо утомления Причинами тонуса могут быть:
• потоки афферентных импульсов, постоянно поступающие с неадаптирующихся рецепторов;
• гуморальные факторы, постоянно присутствующие в плазме крови;
• спонтанная биоэлектрическая активность нейронов (автоматия);
• циркуляция (реверберация) импульсов в ЦНС.
12) Нервный центр состоит из нейронов, а они очень чувствительны к изменению состава внутренней среды организма, что и отражается на свойствах нервных центров. Наиболее важными факторами, влияющими на работу нервных центров, являются: гипоксия; недостаток питательных веществ (например, глюкозы); изменение температуры; воздействие продуктов метаболизма; воздействие различных токсических и фармакологических препаратов.
Разные нервные центры имеют неодинаковую чувствительность к воздействию названных факторов. Так, нейроны коры больших полушарий наиболее чувствительны к гипоксии, недостатку глюкозы, продуктам метаболизма; клетки гипоталамуса – к изменению температуры, содержанию глюкозы, аминокислот, жирных кислот и др.; различные участки ретикулярной формации выключаются разными фармакологическими препаратами, различные нервные центры избирательно активируются или тормозятся разными медиаторами.
13) Пластичность нервного центра означает его способность изменять при определенных обстоятельствах свои функциональные свойства. В основе этого явления лежит поливалентность нейронов нервных центров. Особенно ярко проявляется это свойство при всевозможных повреждениях ЦНС, когда организм компенсирует утраченные функции за счет сохранившихся нервных центров. Особенно хорошо свойство пластичности выражено в коре больших полушарий. Например, центральные параличи, связанные с патологией двигательных центров коры, иногда полностью компенсируются, и ранее утраченные двигательные функции восстанавливаются.
Все основные формы деятельности нервной системы в сложных многоклеточных организмах животных и человека связаны с функционированием определенных групп нервных клеток – нервных центров. Нервный центр является центральным компонентом рефлекторной дуги, в нем происходит переработка информации, вырабатывается программа действия, формируется эталон результата.
Нервный центр – это совокупность нейронов, расположенных на различных уровнях ЦНС и регулирующих сложный рефлекторный процесс или функцию. В нервном центре выделяют следующие отделы: низший, или исполнительный, рабочий и высший, или регуляторный.
Низший (исполнительный) отдел нервного центра локализован в спинном мозге и передает информацию от рабочего отдела к рабочим органам.
Рабочий отдел нервного центра – это отдел, ответственный за осуществление данной функции, как правило, находится в стволовых отделах головного мозга.
Высший (регуляторный) отдел нервного центра расположен в коре больших полушарий мозга и регулирует активность рабочего отдела нервного центра, он вмешивается в регуляцию функций эпизодически, при необходимости корректировки автоматической деятельности рабочего отдела. Высшие отделы включаются в работу нервного центра по условно-рефлекторному механизму. Активность регуляторного (высшего) отдела нервного центра зависит от функционального состояния рабочего отдела.
Строение нервного центра можно рассмотреть на примере дыхательного центра. Исполнительный отдел дыхательного нервного центра расположен в передних рогах грудного отдела спинного мозга и транслирует приказы рабочего центра к дыхательным мышцам. Рабочий отдел представлен центрами вдоха, выдоха и пневмотаксиса, расположенными в продолговатом мозге и варолиевом мосту. Нарушение этого отдела вызывает остановку дыхания. Регуляторный (высший) отдел дыхательного центра находится в лобной доле коры больших полушарий и позволяет произвольно регулировать легочную вентиляцию (глубину и частоту дыхания). Однако эта произвольная регуляция ограничена, она зависит от функциональной активности рабочего отдела и афферентной импульсации, отражающей состояние внутренней среды (в данном случае pH крови, концентрации СО2 и О2 в крови).
Свойства нервных центров обусловлены особенностями проведения нервных импульсов через синапсы, соединяющие различные нервные клетки:
- 1. Одностороннее проведение возбуждения – импульс проводится только в одном направлении, обратное проведение возбуждения через синапс невозможно.
- 2. Наличие латентного периода от начала действия сигнала до проявления рефлекторного акта, так называемая синаптическая задержка. Она обусловлена тем, что на выделение и диффузию медиатора в синапсе требуется промежуток времени в 1,5–2 мс. Соответственно, чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса.
- 3. Суммация возбуждений. В работе нервных центров происходят процессы пространственной и временной суммации подпороговых (недостаточных для передачи импульса через синапс) раздражений. Временная суммация наблюдается, если множество слабых импульсов приходит к нейрону по одному и тому же пути через один синапс с коротким интервалом времени. В результате их действие суммируется, приводя к генерации возбуждения. Пространственная суммация связана с суммированием подпороговых потенциалов, возникающих одновременно в разных синапсах одного нейрона. Оба вида суммации происходят в области аксонного холмика нейрона.
- 4. Более низкая скорость передачи импульса в синапсепо сравнению с передачей его по аксону (около 50–100 импульсов в секунду, что в 5-6 раз ниже скорости передачи в аксоне).
- 5. Утомление нервных центров – длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения. Этот процесс связан с деятельностью синапсов – в них наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, снижается реакция постсинаптического рецептора на медиатор. Разные нервные центры имеют различную скорость утомления. Менее утомляемы центры вегетативной нервной системы, координирующие работу внутренних органов. Значительно более утомляемы центры соматической нервной системы, управляющие произвольной скелетной мускулатурой.
- 6. Трансформация ритма – нервные клетки обладают свойством изменять частоту передающихся импульсов. Характер ответного разряда нейрона зависит от свойств раздражителя, а также и от функционального состояния самого нейрона (его мембранного заряда, возбудимости, лабильности). В обычных условиях – чем сильнее раздражение, тем интенсивнее проявляется ответ.
- 7. В нервных клетках осуществляется интенсивный обмен веществ, для чего необходимо постоянное поступление достаточного количества энергии и кислорода. Особенно чувствительны к недостатку кислорода нервные клетки коры больших полушарий головного мозга: после 5-6 мин кислородного голодания они погибают, даже кратковременное ограничение мозгового кровообращения приводит у человека к потере сознания. Недостаточное снабжение кислородом легче переносят нервные клетки мозгового ствола: их функция восстанавливается через 15–20 мин после полного прекращения кровоснабжения. А функция клеток спинного мозга восстанавливается даже после 30 мин отсутствия кровоснабжения.
- 8. Нервные центры всегда находятся в тонусе, который обеспечивается постоянно поступающими импульсами от разных структур мозга и исполнительных органов. В ответ центры посылают редкие импульсы к органам, поддерживая в них соответствующий тонус. Даже во время сна мышцы не расслабляются полностью и контролируются соответствующими центрами.
- 9. Нервные центры чувствительны к химическим веществам (в том числе лекарственным), проникающим через гематоэицефалический барьер, и обладают специфической реакцией на различные вещества. К примеру, стрихнин возбуждает нервные центры, блокируя работу тормозных синапсов; хлороформ и эфир сначала возбуждают, а затем подавляют работу нервных центров; апоморфин возбуждает рвотный центр; цититои и лобелии возбуждают дыхательный центр; морфин его угнетает, коразол возбуждает клетки двигательной зоны коры, вызывая судороги.
- 10. После окончания действия раздражителя активное состояние нервного центра продолжается еще некоторое время – так называемое последействие, или следовые процессы. Длительность следовых процессов различна: в спинном мозге – несколько секунд или минут, в подкорковых центрах мозга – десятки минут, часы и даже дни, в коре больших полушарий могут сохраняться до нескольких десятков лет. Следовые процессы имеют важное значение в понимании механизмов памяти. Непродолжительное последействие до 1 ч связано с циркуляцией (реверберацией) импульсов в нервных цепях (Р. Лоренте де Но, 1934) и обеспечивает согласно ревербационной теории кратковременную память. Согласно биохимической теории памяти (X. Хиден, 1969) механизмы долговременной памяти обеспечиваются изменением структуры белков нейронов и глиальных клеток: при запоминании происходят структурные изменения в молекулах РНК, на основе которых строятся новые белки, несущие информацию о прежних раздражителях. Эти белки длительно содержатся в нейронах, а также в глиальных клетках головного мозга.
Читайте также:
