
Структурная и функциональная характеристика нервных волокон
Структурно-функциональная характеристика нервных волокон. Нервные волокна — это отростки нейронов, с помощью которых осуществляется связь между нейронами и с исполнительными клетками. Имеется два типа нервных волокон: миелинизированные и немиелинизированные (безмиелиновые). Оболочку безмиелиновых волокон образуют шванновские клетки (леммоциты), в которые погружаются осевые цилиндры нервных волокон. Оболочку миелинизированных волокон образуют в периферической нервной системе также шванновские клетки (миелоциты, глиальная клетка), формирующие миелин (многослойная обертка мембран миелоцитов — до 100), а в ЦНС — олигодендроциты (глиальные клетки, т.е. те же шванновские клетки). Миелиновая оболочка через равные участки (около 1 мм) прерывается, образуя свободные от миелина небольшие участки — перехваты Ранвье. Основную часть миелина (78% сухого веса) составляют липиды, обеспечивающие изолирующие свойства оболочки. Нервные волокна обеспечивают проведение возбуждения и аксонныйтра транспорт, выполняющий трофическую функцию нейрона.
Выделяют три типа волокон: А, В и С (см. табл. 3).
Таблица 3. Типы волокон в нервах млекопитающих (по Эрлангеру-Гассеру)
| Тип волокон | Диаметр волокна, мкм | Скорость проведения возбуждения, м/с | Длительность абсолютного рефрактерного периода, мс |
| Аα (миел.) | 12-20 | 70-120 | 0,4-1,0 |
| Аβ (миел.) | 5-12 | 30-70 | - |
| Аσ (миел.) | 3-6 | 15-30 | 0.4-1,0 |
| Аγ (миел.) | 2-5 | 12-30 | - |
| В (миел.) | 1-3 | 5-12 | 1,2 |
| С (немиел.) | 0,3-1,3 | 0,5-2,3 |
Волокна типов А и В являются миелинизированными. А-волокна — это афферентные и эфферентные волокна соматической нервной системы; к волокнам типа В принадлежат преганглионарные волокна ВНС. С-волокна немиелинизированные — это постганглионарные волокна ВНС, а также афферентные волокна от некоторых болевых, тепловых и висцеральных рецепторов.
Аксонный транспорт. Основная масса веществ образуется в трофическом центре нейрона, расположенном преимущественно около ядра, а используются эти вещества в различных участках тела нейрона и его отростках. В аксонных окончаниях также происходит синтез медиаторов, АТФ и повторное использование мембраны пузырьков после высвобождение медиатора. Выделяют быстрый и медленный аксонный транспорт (оба они с затратой энергии).
Быстрый аксонный транспорт осуществляется со скоростью 200—400 мм/сут от тела клетки до аксонных окончаний — прямой (антероградный) — и в противоположном направлении — обратный (ретроградный) транспорт. Вещества транспортируются с помощью микротрубочек и микрофиламентов, часть которых представляет собой актиновые нити (актин составляет 10—15 % белков нейрона). Посредством прямого транспорта в аксонные окончания доставляются митохондрии, ферменты, медиаторы, липиды, везикулы, содержащие гликопротеиды мембран, специальные белки и пептиды (нейротрофогены). Посредством обратного транспорта в тело нейрона переносятся везикулы, содержащие остатки разрушенных структур, фрагменты мембран, факторы роста нервов и другие ростовые факторы, регулирующие синтез белка в соме клетки. Многие вещества, доставленные посредством ретроградного транспорта, подвергаются разрушению в лизосомах. В патологических случаях по аксону к телу клетки могут транспортироваться столбнячный экзотоксин, вирусы полиомиелита, герпеса, бешенства.
Медленный аксонный транспорт идет в прямом направлении и представляет собой передвижение всего столба аксоплазмы со скоростью 1—2 мм/сут. С помощью этого транспорта перемещаются образованные в эндоплазматической сети белки микротрубочек и микрофиламентов (актин, тубулин и др.), ферменты цитозоля, РНК, белки каналов, насосов и другие вещества.
Значение аксонного транспорта: 1) необходим для поддержания структуры нервного волокна; 2) необходим для аксонного роста и образования синаптических контактов; 3) играет важную роль при регенерации нервных волокон. На мышечное волокно такое влияние оказывают нейротрофогены (специальные белки, β-эндорфин и другие пептиды); обратное влияние на мотонейрон осуществляется с помощью миотрофогенов (фактора роста нервов, инсулиноподобного фактора роста).
Результат повреждения нервного волокна. Если нервное волокно в результате травмы разорвано, его периферический отрезок, лишенный связи с телом нейрона, подвергается разрушению, которое называется валлеровской дегенерацией. В течение 2-3 сут наступает распад нейрофибрилл, митохондрий, миелина и синаптических окончаний. Участок волокна, связанный с телом нейрона, регенерирует, проводимость восстанавливается. В норме нервное волокно, как и нейрон, функционирует на протяжении всей жизни организма и проводит возбуждение без декремента (затухания) по всей длине волокна, например от тела мотонейронов спинного мозга до мышечных волокон конечностей (до 1 м).
Механизм проведения возбуждения. Проведение ПД возможно только при наличии на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциалзависимых ионных каналов, ответственных за формирование новых ПД. Это осуществляется, согласно нашим представлениям, следующим образом. В распространении ПД можно выделить два этапа: этап распространения электрического поля, снижающего мембранный потенциал, и этап генерации новых ПД в новых участках нервного волокна. Электрическое поле — разновидность материи, посредством которой осуществляется силовое воздействие на электрические заряды, находящиеся в этом поле. Электрическое поле, которое генерируется биологическими структурами, является источником информации о состоянии клеток и органов организма. Например, состояние электрического поля сердца, записанного в виде электрокардиограммы, помогает выявить его возможные повреждения. В зависимости от расположения и концентрации ионных каналов, в мембране нервного или мышечного волокна имеется два варианта проведения ПД: непрерывное и сальтаторное (скачкообразное).
Непрерывное проведение ПД происходит в мышечных волокнах и в безмиелиновых нервных волокнах (тип С), имеющих равномерное распределение потенциалуправляемых ионных каналов по всей длине волокна. Проведение нервного импульса начинается (как и в мышечном волокне) с распространения колеблющегося по величине электрического поля. Амплитуда ПД в нервном волокне (мембранный потенциал + инверсия) составляет 100-120 мВ, постоянная длины мембраны (λm— расстояние, на котором сохраняется 37 % величины ПД в виде электрического поля) в безмиелиновых волокнах равна 0,1—1,0 мм. В связи с этим возникший ПД за счет действия своего электрического поля способен деполяризовать мембрану соседнего участка до критического уровня на расстояние от 0,1 до 1,0 мм. Это означает, что на этом участке (0,1-1,0 мм) одновременно генерируются новые ПД, обусловленные движением ионов Na + в клетку, К + — из клетки (на распространение электрического поля время не затрачивается). Число одновременно возникающих ПД ограничивается длиной возбужденного участка — для безмиелинового волокна 0,1-1,0 мм (ПД возникают рядом друг с другом в непосредственной близости). Причем сами ПД не перемещаются (они исчезают там, где возникают). Главную роль в возникновении новых ПД играет передний ПД. Вспомогательную роль в генерации новых ПД в невозбужденных участках нервного волокна играют промежуточные ПД (возникшие сзади переднего ПД) — их электрическое поле суммируется с электрическим полем переднего ПД, но они находятся дальше от участка нервного волокна, где возникают новые ПД. Таким образом, непрерывное распространение нервного импульса, как и сальтаторное (см. ниже), идет через генерацию новых ПД по эстафете, когда каждый участок мембраны сначала выступает как раздражаемый электрическим полем, а затем — как раздражающий (в результате формирования в нем новых ПД) (рис. 15).
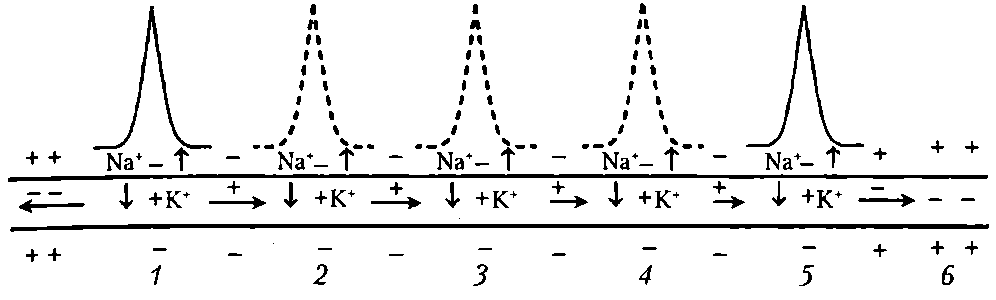
Рис. 15. Непрерывное проведение возбуждения (ПД) в безмиелиновом нервном волокне. Уменьшение длины горизонтальных стрелок иллюстрирует ослабление электрического поля переднего ПД, инициирующего возбуждение соседнего участка волокна: 1-5 - состояние возбуждения (ПД); 6 – состояние покоя; пунктиром обозначены промежуточные ПД; вертикальные стрелки указывают направление движения Na + в клетку и K + - из клетки
Сальтаторное проведение ПД происходит по миелинизированным волокнам (типы А и В), для которых характерна концентрация потенциалуправляемых ионных каналов только в небольших участках мембраны (в перехватах Ранвье), где их плотность достигает 10 тыс. на 1 мкм 2 , что примерно в 100 раз выше, чем в любом участке мембраны безмиелиновых волокон. В области миелиновых муфт, обладающих хорошими изолирующими свойствами, потенциалуправляемых каналов почти нет, поэтому ПД здесь не возникают. ПД, возникший в одном перехвате Ранвье, за счет действия своего электрического поля деполяризует мембрану соседних перехватов до КП, что приводит к возникновению в них новых ПД, т.е. возбуждение возникает как бы скачкообразно — только в перехватах (рис.16).
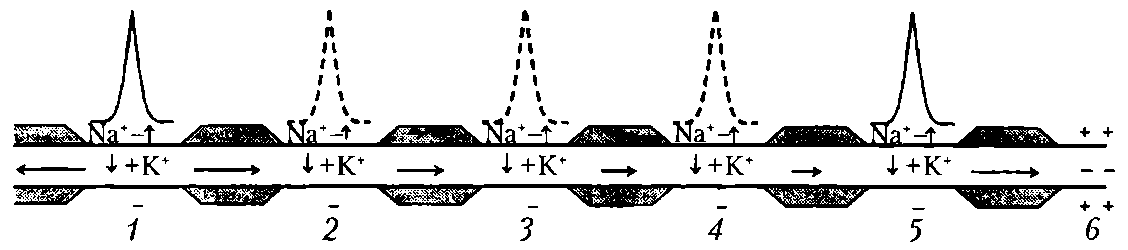
Рис. 16. Сальтаторное проведение возбуждения (ПД) в миелинизированном нервном волокне. Уменьшение длины горизонтальных стрелок иллюстрирует ослабление электрического поля переднего ПД, инициирующего возбуждение соседнего участка волокна: 1-5 - состояние возбуждения (ПД); 6 – состояние покоя; пунктиром обозначены промежуточные ПД; вертикальные стрелки указывают направление движения Na + в клетку и K + - из клетки
Напомним, что Nа-каналы начинают открываться при достижении деполяризации клеточной мембраны 50% КП. Постоянная длины мембраны миелинового волокна достигает 5 мм. Это означает, что электрическое поле ПД на данном расстоянии сохраняет 37% своей амплитуды (около 30 мВ) и может деполяризовать мембрану до КП (ΔV в перехватах Ранвье составляет около 15 мВ). Благодаря этому, в случае повреждения ближайших на пути следования перехватов Ранвье ПД возбуждает 2—4-й и даже 5-й перехваты. Поэтому возбуждение распространяется очень быстро по всей длине волокна, а ионы движутся только перпендикулярно относительно длины волокна — в клетку и из клетки (вдоль волокна они не успевают смещаться). Электрическое поле ПД, возникших сзади переднего (промежуточные ПД), суммируется с электрическим полем переднего ПД, как и при непрерывном распространении возбуждения.
Простые расчеты показывают, что скорость проведения ПД по нервному волокну за счет движения ионов вдоль волокна была бы слишком низкой. В частности, скорость движения Na + в клетку согласно электрохимическому градиенту легко рассчитать по толщине клеточной мембраны (6—10 нм) и длительности ПД (например, толстого миелинизированного волокна — около 1 мс) — вход Na + в клетку и выход К + из клетки. При этом Na + при движении в клетку согласно электрохимическому градиенту преодолевает расстояние в 8 нм примерно за 0,5 мс (восходящая часть пика ПД продолжается около 0,5 мс). На основании этого рассчитаем, сколько времени потребуется на прохождение ПД 1 м. Для прохождения ионом Na + 8 нм необходимо 0,5 мс.
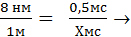
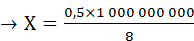
≈17 часов,
т.е. возбуждение распространилось бы на 1 мм за 17 часов.
Легко представить, какова была бы подвижность (точнее, неподвижность) живых существ, если бы возбуждение вдоль нервных и мышечных волокон распространялось со скоростью диффузии ионов, но очень трудно представить, как бы выглядел весь животный мир! Чтобы шевельнуть пальцем пришлось бы ждать около суток!
Возникающие ПД не могут инициировать развитие других ПД в обратном направлении, так как нервное волокно находится еще в рефрактерном состоянии. Это не противоречит тому факту, что раздражение нервного волокна в эксперименте вызывает распространение возбуждения в двух направлениях, поскольку в этом случае участки нервного волокна по обеим сторонам от места раздражения находятся в состоянии покоя. В натуральных же условиях первый ПД, возникший на мембране тела нейрона, инициирует распространение возбуждения только в одном направлении — по аксону к другой клетке.
Сравнение механизма непрерывного и сальтаторного проведения возбуждения показывает, что различие между ними не принципиально. Оно заключается лишь в том, что очередные ПД в безмякотном волокне возникают на более близком расстоянии друг от друга, поскольку ионные каналы расположены в непосредственной близости друг от друга и непрерывно по всей длине нервного волокна. Поэтому такое проведение и назвали непрерывным.
Однако сальтаторное проведение возбуждения имеет два важных преимущества перед непрерывным. Во-первых, сальтаторное проведение более экономично с точки зрения затрат энергии, так как возбуждаются только перехваты Ранвье, площадь которых менее 1% мембраны, и, следовательно, требуется меньше энергии для восстановления трансмембранных градиентов Na + и К + , расходующихся в процессе возникновения ПД (в миелинизированных волокнах при распространении возбуждения теряется ионов в 100 раз меньше, чем в немиелинизированных). Во-вторых, возбуждение в миелинизированных волокнах проводится с большей скоростью (см. таблицу), чем в безмиелиновых волокнах, так как в них электрическое поле ПД распространяется значительно дальше на соседние перехваты Ранвье, поскольку электроизоляция (миелиновые муфты) уменьшает рассеивание электрического поля.
В процессе проведения возбуждения время затрачивается только на перпендикулярное относительно мембраны волокна движение ионов в клетку и из клетки при формировании нового ПД, а влияние электрического поля возникших ПД на соседний участок распространяется вдоль длины волокна мгновенно — время распространения электрического поля практически равно нулю. Скорость распространения возбуждения увеличивается также при большой амплитуде ПД, что является следствием формирования более сильного электрического поля, обеспечивающего критический уровень деполяризации нервного волокна на большем расстоянии и быстрее.
В настоящее же время одни физиологи считают, что возникший ПД электротонически вызывает возбуждение в соседнем участке нервного или мышечного волокна; по мнению других, W. Willis (2004) и R. Klinke (2004), это осуществляется с помощью локальных входящих и выходящих токов ионов и между соседними участками волокна в продольном направлении. Однако электротон — это частичная деполяризация мембраны, еще не активирующая ионные каналы, поэтому в первом случае возбуждение вообще не возникло бы, а во втором случае оно распространялось бы слишком медленно.
Скорость распространения электрического поля хорошо иллюстрируется процессом общения по телефону — собеседники могут находиться на расстоянии нескольких тысяч километров друг от друга, но слова они слышат через такой же промежуток времени, как и при непосредственном разговоре.
Характеристика проведения возбуждения по нервному волокну. 1. Двустороннее проведение возбуждения можно продемонстрировать в эксперименте при нанесении раздражения в любом участке нерва или нервного волокна, при этом возбуждение регистрируется как в одном, так и в другом направлении от места раздражения.
2. Изолированное проведение возбуждения в отдельных волокнах нервного ствола обусловлено тем, что влияние электрического поля ПД соседнего волокна не возбуждает другие волокна нерва вследствие изолирующего эффекта их оболочек и интерстиция. Изолированное проведение импульсов по нервным волокнам обеспечивает точное афферентное и эфферентное влияния функционально разнородных волокон нерва. Однако если одновременно возбуждается значительное количество волокон, то возникает достаточно сильное электрическое поле, способное открыть ворота натриевых каналов соседних и таким образом усилить нервное влияние на эффекторные клетки или нейроны в регуляторных процессах.
3. Бездекрементное (без затухания; лат. decrementum — убывание, уменьшение) проведение по всей длине нервного волокна, так как ПД возникают в каждом участке волокна заново под влиянием предыдущего ПД, и его величина в каждом участке волокна складывается из величины ПП и фазы инверсии ПД.
4.Большая скорость проведения возбуждения (до 120 м/с) в нервных волокнах типа Аα. Для сравнения отметим, что скорость передачи гуморальных влияний ограничена скоростью кровотока — от 0,5 мм/с в капиллярах до 0,25 м/с в аорте (время полного кругооборота крови — около 22 с). Большая скорость распространения ПД обеспечивает быстрое влияние на другие нейроны, рабочие органы, получение обратной информации, играющей важную роль в регуляторных процессах.
Малая утомляемость нервного волокна впервые была показана Н.Е. Введенским (1883): в проводимых опытах нерв сохранял способность к проведению возбуждения в течение 6—8 ч непрерывного раздражения несильными импульсами тока при условии наличия кислорода в окружающей среде и поддержания влажного состояния нерва. Это обусловлено тем, что при проведении ПД по нервным волокнам используется незначительная часть запасов трансмембранных ионных градиентов и, следовательно, нужны небольшие количества АТФ для их восстановления. Расход энергии в нервном волокне на единицу массы в миллион раз меньше, чем в работающей мышце.
Высокая лабильность — нервное волокно может проводить до 500—1000 имп./с.
Возбуждение от нервного волокна передается к другой клетке с помощью синапса.
Нервное волокно - аксон - покрыт клеточной мембраной.
Выделяют 2 вида нервных волокон.
Безмиелиновые нервные волокна- один слой швановских клеток, между ними - щелевидные пространства. Клеточная мембрана на всем протяжении контактирует с окружающей средой. При нанесении раздражения возбуждение возникает в месте действия раздражителя. Безмиелиновые нервные волокна обладают электрогенными свойствами (способностью генерировать нервные импульсы) на всем протяжении.
Миелиновые нервные волокна- покрыты слоями шванновских клеток, которые местами образуют перехваты Ранвье (участки без миелина) через каждые 1 мм. Продолжительность перехвата Ранвье 1 мкм. Миелиновая оболочка выполняет трофическую и изолирующую функции (высокое сопротивление). Участки, покрытые миелином не обладают электрогенными свойствами. Ими обладают перехваты Ранвье. Возбуждение возникает в ближайшем к месту действия раздражителя перехвата Ранвье. В перехватах Ранвье высокая плотность Nа-каналов, поэтому в каждом перехвате Ранвье происходит усиление нервных импульсов.
Перехваты Ранвье выполняют функцию ретрансляторов (генерируют и усиливают нервные импульсы).
Механизм проведения возбуждения по нервному волокну
1885 г. - Л. Герман - между возбужденными и невозбужденными участками нервного волокна возникают круговые токи.
При действии раздражителя имеется разность потенциалов между наружной и внутренней поверхностями ткани (участки несущие различные заряды). Между этими участками возникает электрический ток (движение ионов Nа+). Внутри нервного волокна возникает ток от положительного полюса к отрицательному полюсу, т. е. ток направлен от возбужденного участка к невозбужденному. Этот ток выходит через невозбужденный участок и вызывает его перезарядку. На наружной поверхности нервного волокна ток идет от невозбужденного участка к возбужденному. Этот ток не изменяет состояние возбужденного участка, т. к. он находится в состоянии рефрактерности.
Доказательство наличия круговых токов:нервное волокно помещают в раствор NaCl и регистрируют скорость проведения возбуждения. Затем нервное волокно помещают в масло (повышается сопротивление) - скорость проведения уменьшается на 30 %. После этого нервное волокно оставляют на воздухе - скорость проведения возбуждения уменьшается на 50 %.
Особенности проведения возбуждения по миелиновым и безмиелиновым нервным волокнам:
1. миелиновые волокна- имеют оболочку обладающую высоким сопротивлением, электрогенные свойства только в перехватах Ранвье. Под действием раздражителя возбуждение возникает в ближайшем перехвате Ранвье. Соседний перехват в состоянии поляризации. Возникающий ток вызывает деполяризацию соседнего перехвата. В перехватах Ранвье высокая плотность Nа-каналов, поэтому в каждом следующем перехвате возникает чуть больший (по амплитуде) потенциал действия, за счет этого возбуждение распространяется без декремента и может перескакивать через несколько перехватов. Это сальтаторная теория Тасаки. Доказательство теории - в нервное волокно вводили препараты, блокирующие несколько перехватов, но проведение возбуждения регистрировалось и после этого. Это высоко надежный и выгодный способ, т. к. устраняются небольшие повреждения, увеличивается скорость проведения возбуждения, уменьшаются энергетические затраты;
2. безмиелиновые волокна- поверхность обладает электрогенными свойствами на всем протяжении. Поэтому малые круговые токи возникают на расстоянии в несколько микрометров. Возбуждение имеет вид постоянно бегущей волны.
Этот способ менее выгоден: большие затраты энергии (на работу Nа-К-насоса), меньшая скорость проведения возбуждения.
Классификация нервных волокон
Нервные волокна классифицируются по:
1. длительности потенциала действия;
2. строению (диаметру) волокна;
3. скорости проведения возбуждения.
Выделяют следующие группы нервных волокон:
1. группа А (альфа, бета, гамма, дельта) - самый короткий потенциал действия, самая толстая миелиновая оболочка, самая высокая скорость проведения возбуждения;
2. группа В - миелиновая оболочка менее выражена;
3. группа С - без миелиновой оболочки.
Синапсы ЦНС
Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку).
Cтруктура синапса:
1) пресинаптическая мембрана (электрогенная мембрана в терминале аксона, образует синапс на мышечной клетке);2) постсинаптическая мембрана (электрогенная мембрана иннервируемой клетки, на которой образован синапс);3) синаптическая щель (пространство между пресинаптической и постсинаптической мембраной, заполнена жидкостью, которая по составу напоминает плазму крови).
2) Существует несколько классификаций синапсов.
1) центральные синапсы;2) периферические синапсы.
Центральные синапсы лежат в пределах центральной нервной системы, а также находятся в ганглиях вегетативной нервной системы. Центральные синапсы – это контакты между двумя нервными клетками, причем эти контакты неоднородны и в зависимости от того, на какой структуре первый нейрон образует синапс со вторым нейроном, различают:
1) аксосоматический, образованный аксоном одного нейрона и телом другого нейрона;2) аксодендритный, образованный аксоном одного нейрона и дендритом другого;3) аксоаксональный (аксон первого нейрона образует синапс на аксоне второго нейрона);4) дендродентритный (дендрит первого нейрона образует синапс на дендрите второго нейрона).
Различают несколько видов периферических синапсов:
1) мионевральный (нервно-мышечный), образованный аксоном мотонейрона и мышечной клеткой;2) нервно-эпителиальный, образованный аксоном нейрона и секреторной клеткой.
Функциональная классификация синапсов:1) возбуждающие синапсы;2) тормозящие синапсы.
По механизмам передачи возбуждения в синапсах:1) химические;2) электрические.
Особенность химических синапсов заключается в том, что передача возбуждения осуществляется при помощи особой группы химических веществ – медиаторов.
Различают несколько видов химических синапсов:1) холинэргические. В них происходит передача возбуждения при помощи ацетилхолина;2) адренэргические. В них происходит передача возбуждения при помощи трех катехоламинов;3) дофаминэргические. В них происходит передача возбуждения при помощи дофамина;4) гистаминэргические. В них происходит передача возбуждения при помощи гистамина;5) ГАМКэргические. В них происходит передача возбуждения при помощи гаммааминомасляной кислоты, т. е. развивается процесс торможения.
Особенность электрических синапсов заключается в том, что передача возбуждения осуществляется при помощи электрического тока. Таких синапсов в организме обнаружено мало.
Синапсы имеют ряд физиологических свойств:1) клапанное свойство синапсов, т. е. способность передавать возбуждение только в одном направлении с пресинаптической мембраны на постсинаптическую;2) свойство синаптической задержки, связанное с тем, что скорость передачи возбуждения снижается;3) свойство потенциации (каждый последующий импульс будет проводиться с меньшей постсинаптической задержкой). Это связано с тем, что на пресинаптической и постсинаптической мембране остается медиатор от проведения предыдущего импульса;4) низкая лабильность синапса (100–150 имульсов в секунду).
Скорость проведения возбуждения через синапс намного меньше, чем по нервному волокну, так как здесь тратится время на активацию пресинаптической мембраны, переход через нее кальция, выделение ацетилхолина в синаптическую щель, деполяризацию постсинаптической мембраны, развитие ПКП.Синаптическая передача возбуждения имеет рад свойств:

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.
Отростки нервных клеток, покрытые оболочками, называются нервными волокнами. В соответствии с особенностями их строения все нервные волокна делятся на две основные группы — миелиновые и безмиелиновые. Те и другие состоят из отростка нервной клетки, который лежит в центре волокна и поэтому называется осевым цилиндром, и оболочки, образованной клетками олигодендроглии, которые здесь называются нейролеммоцитами(шванновскими клетками).
Безмиелиновые нервные волокна находятся преимущественно в составе вегетативной нервной системы.
Клетки олигодендроглии оболочек безмиелиновых нервных волокон. Располагаясь плотно, образуют тяжи, в которых на определенном расстоянии друг от друга видны овальные ядра. В нервных волокнах внутренних органов, как правило, в таком тяже располагается не один, а несколько (10—20) осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в смежное, такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. При электронной микроскопии безмиелиновых нервных волокон видно, что по мере погружения осевых цилиндров в тяж леммоцитов последние одевают их как муфтой. Оболочки леммоцитов при этом прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану — мезаксон, на которой как бы подвешен осевой цилиндр. Оболочки нейролеммоцитов очень тонкие, поэтому ни мезаксона, ни границ этих клеток под световым микроскопом нельзя рассмотреть, и оболочка безмиелиновых нервных волокон в этих условиях выявляется как однородный тяж цитоплазмы, "одевающий" осевые цилиндры. С поверхности каждое нервное волокно покрыто базальной мембраной.
Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Диаметр поперечного сечения их колеблется от 1 до 20 мкм. Они также состоят из осевого цилиндра, "одетого" оболочкой из нейролеммоцитов, но диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее. В сформированном миелиновом волокне принято различать два слоя оболочки: внутренний, более толстый, миелиновый слой и наружный, тонкий, состоящий из цитоплазмы и ядер нейролеммоцитов — нейролемму.
Миелиновое нервное волокно представляется однородным цилиндром, в котором на определенном расстоянии друг от друга располагаются светлые линии-насечки миелина. Через некоторые интервалы встречаются участки волокна, лишенные миелинового слоя — узловые перехваты — перехваты Ранвье.
Осевой цилиндр нервных волокон состоит из нейроплазмы—цитоплазмы нервной клетки, содержащей продольно ориентированные нейрофиламенты и нейротубулы. В нейроплазме осевого цилиндра лежат митохондрии, которых больше в непосредственной близости к перехватам и особенно много в концевых аппаратах волокон. С поверхности осевой цилиндр покрыт мембраной — аксолеммой, обеспечивающей проведение нервного импульса. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Тонкие волокна, бедные миелином и безмиелиновые волокна проводят нервный импульс со скоростью 1—2 м/с, тогда как толстые миелиновые волокнасо скоростью 5—120 м/с.
В безмиелиновом волокне волна деполяризации мембраны идет по всей плазмолемме, не прерываясь, а в миелиновом волокне возникает только в области перехвата. Таким образом, для миелиновых волокон характерно сальтаторное проведение возбуждения, то есть прыжками. Между перехватами по аксолемме идет электрический ток, скорость которого выше, чем прохождение волны деполяризации.
Читайте также:
