
Структуры цнс обеспечивающие дыхательную периодику
Тема занятия: Механизмы ритмогенеза в дыхательном центре. Регуляция дыхания в разных экспериментальных условиях. Функциональная система поддержания газового гомеостаза крови
Мотивационная характеристика темы.Важнейшая функция регуляции дыхания – поддержание постоянства показателей внутренней среды организма: рО2 , рСО2 , рН крови. Нарушение регуляции дыхания часто приводит к необратимым изменениям в организме и служит причиной его гибели. Знание механизмов, обеспечивающих регуляцию акта дыхания в физиологических условиях, позволяет дать правильную оценку эффективности дыхания на разных его этапах в условиях нормы и патологии.
Цель занятия: 1.Изучить механизмы саморегуляции дыхания человека. 2. Изучить функциональную систему поддержания газового состава крови. 3. Изучить особенности регуляции дыхания при физической нагрузке, при изменении атмосферного давления.
Конкретные цели
| Знать | Уметь |
| 1. Структуры ЦНС, обеспечивающие дыхательную периодику. 2. Рецепторы легких, их роль в регуляции дыхания. 3. Значение проприорецепторов мышц в регуляции дыхания. 4. Роль гипоталамуса, лимбической системы, коры больших полушарий в регуляции дыхания. 5. Влияние на частоту и глубину дыхания газового состава и рН крови, периферические и центральные хеморецепторы. 6. Функциональную систему обеспечения газового гомеостаза. 7. Дыхание при физической нагрузке, при изменении барометрического давления. 8. Особенности первого вдоха новорожденного. 9. Возрастные особенности регуляции дыхания. | 1. Объяснять принцип и сущность метода пневмографии. 2. Проводить самостоятельно под контролем преподавателя: -пробу с физической нагрузкой, - пробу Штанге-Генча - анализировать по пневмограмме ритм дыхания 3. Рисовать и объяснять рефлекторные дуги безусловных рефлексов, обеспечивающих регуляцию дыхания и ритмогенез в дыхательном центре. 4. Использовать полученные знания для решения учебных задач. |
Вопросы, изученные ранее и необходимые для освоения данной темы
1. Строение и физиология скелетных мышц.
2. Строение и физиология ЦНС.
3. Иннервация скелетных мышц: расположение альфа-мотонейронов спинного мозга, роль супраспинальных структур.
4. Роль проприорецепторов в регуляции тонуса скелетных мышц.
5. Расположение вегетативных центров; влияние симпатической и парасимпатической нервной системы на диаметр бронхов, секрецию желез, тонус нервных центров.
Содержание обучения
I. Рекомендуемая учебная литература:
1. Лекционный материал.
2. Физиология человека. Под ред. Г.И. Косицкого, М. , 1985., с. 304-319.
3. Физиология человека. Под ред. Б.И. Ткаченко, СПб 1994, с. 362-378.
4. Физиология человека. Под ред. Б.М. Покровского, Г.Ф. Коротько, том 1., с. 422-442.
5. Физиология Человека. Под ред. Н.А. Агаджаняна, В.И. Циркина, М. , 2001., с. 279- 284, с. 292-294.
6. Обучающая программа по физиологии дыхания.
II. Литература дополнительная:
2. Филимонов В.И. Руководство по общей и клинической физиологии. М. 2002. с. 628-641.
3. Физиология человека под ред. Шмидта и Тевса. М., 1989. Том 3, С. 594-603, 635-640.
III. Письменно в тетради для протоколов изобразить и объяснить схемы:
ІV. Для контроля усвоения знаний ответить на следующие вопросы:
1. Общий принцип регуляции внешнего дыхания. Характеристика основных регулируемых констант: рО2 , рСО2 в артериальной крови. Типы легочного дыхания (эупноэ, тахипноэ, брадипноэ, апноэ; нормо-, гипо- и гипервентиляция).
2. Дыхательный центр, его локализация. Экспериментальное доказательство его локализации (изменение дыхания при пересечении мозга на разных уровнях).
3. Локализация нейронов, иннервирующих дыхательные мышцы в спинном мозге.
4. Бульбарный центр (центр судорожного дыхания), его локализация, нейронная структура, роль в регуляции дыхания, автоматия нейронов.
5. Апнеистический и пневмотаксический центры, функции.
6. Роль гипоталамуса, лимбической системы, коры больших полушарий в регуляции дыхания.
7. Рефлекторная регуляция. Виды и локализация рецепторов. Значение газового состава крови для регуляции дыхания. Эксперимент Фредерика. Хеморецепторы.
8. Особенности первого вдоха новорожденного. Возрастные особенности регуляции дыхания.
9. Значение рецепторов дыхательных путей: рецепторы растяжения легких (рефлекс Геринга-Брейера), ирритантные рецепторы, юкстаальвеолярные рецепторы.
10. Условнорефлекторная регуляция. Регуляция дыхания при физической нагрузке.
11. Функциональная система поддержания газового состава крови. Кратковременные и долговременные адаптационные изменения в работе ФУС при изменении атмосферного давления. Дыхание в горах, дыхание под водой.
Рисунок 3. Схема регуляции дыхания
Рисунок 4. Влияние на дыхание перерезок центральной нервной системы на различных уровнях:
К—представительство дыхательного центра в коре (условно),
Гт — представительство дыхательного центра в гипоталамусе,
П — пневмотаксический центр, Ап — апнеистический центр (варолиев мост),
Э — экспираторный центр (продолговатый мозг),
И — инспираторный центр (продолговатый мозг), С – шейный отдел спинного мозга, Тh – грудной отдел спинного мозга.
а — д — уровни перерезок и соответствующие им пневмограммы (необходимо указать уровень перерезки и тип изменения дыхания)
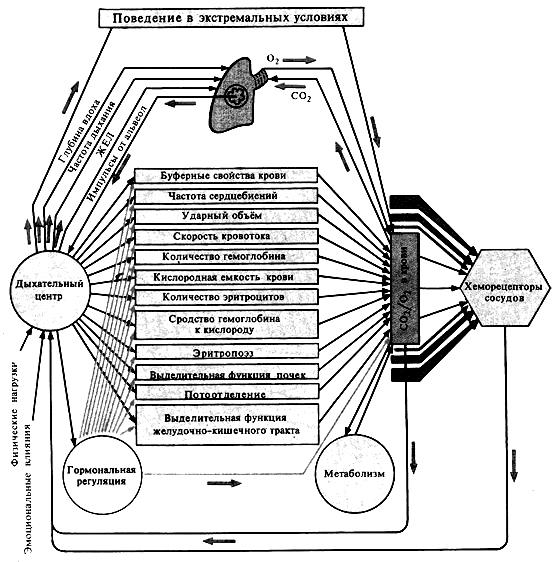
Рисунок 5. Функциональная система, поддерживающая газовый состав внутренней среды организма.
Регуляция дыхания и другие функции организма
Дыхательный центр продолговатого мозга обеспечивает такую вентиляцию легких, которая необходима для поддержания на оптимальном уровне напряжения кислорода и углекислого газа. Напряжение этих газов, воздействуя на дыхательный центр через хеморецепторы, вызывает ответную реакцию дыхания, направленную на устранение отклонения в концентрации. Таким образом осуществляется регуляция дыхания по принципу отклонения регулируемого параметра от нормальных значений.
Вместе с тем, изменения вентиляции легких наблюдаются при самых разнообразных ситуациях, когда нет изменения напряжения кислорода и углекислого газа. Например, воздействия холода или тепла на кожу приводят к возбуждению дыхательного центра и учащению дыхания. Кроме того, на дыхание влияет изменение температуры тела: и понижение, и незначительное повышение вызывают увеличение вентиляции легких. Весьма существенно увеличивает частоту дыхания боль. Вызывают изменения дыхания и физическая, и эмоциональная нагрузки. Такое изменение паттерна дыхания, не обусловленное изменением концентрации газов в крови, является проявлением варианта регуляции по принципу возмущения. Это означает, что тем сигналом, который поступает в дыхательный центр и вызывает гипервентиляцию, служит не отклонение в газовом составе крови, а сигнал о происходящем в организме возмущении, способном привести к отклонению в газовом составе крови, регуляция осуществляется до того, как произойдут отклонения. Механизмы возмущающих влияний на дыхательный центр становятся понятны, если вспомнить о том, что дыхательный центр входит в состав ретикулярной формации ствола мозга.
Ретикулярная формация ствола мозга
Под ретикулярной формацией обычно понимают клеточную массу, лежащую в толще мозгового ствола от нижних отделов продолговатого до промежуточного мозга. Эта клеточная масса слабо структурирована и не имеет четких границ. Внутри ретикулярной формации расположены чувствительные и двигательные ядра продолговатого, среднего и промежуточного мозга. Нейроны ретикулярной формации характеризуются немногочисленными длинными и мало ветвящимися дендритами, их шипики слабо дифференцированы. В медиальной части ретикулярной формации расположены крупные и гигантские клетки, в продолговатом мозге они сконцентрированы в гигантоклеточном ядре. Именно от этих клеток отходят аксоны, которые формируют эфферентные пути. В частности, ретикулоспинальный тракт, пути к таламусу, мозжечку, базальным ганглиям и коре больших полушарий.
Сетевое строение ретикулярной формации обеспечивает высокую надежность ее функционирования и устойчивость к повреждающим воздействиям, потому что локальные повреждения всегда компенсируются за счет сохранившихся элементов сети. Такое сетевое строение обеспечивает и еще одну важную особенность функционирования ретикулярной формации: раздражение любой из ее частей за счет многочисленных связей охватывает всю данную структуру. Кроме того, эффекты стимуляции, как правило, оказываются весьма длительными за счет свойства нейронной сети пролонгировать возбуждение.
Гипоталамус
Существенную роль в регуляции дыхания играет гипоталамус. Основная роль гипоталамуса в изменении паттерна дыхания заключается в том, что ритм дыхания приводится в соответствие с уровнем обмена веществ. Одной из основных особенностей нейронов гипоталамуся является их чувствительность к составу омывающей их крови. Нейроны срединной группы ядер гипоталамуса обладают детектирующей функцией, они реагируют на изменение температуры крови, осмотическую концентрацию, концентрацию глюкозы и аминокислот. Таким образом, в гипоталамус поступает информация о состоянии внутренней среды организма. В гипоталамусе на основании этой информации происходит формирование мотиваций - побуждений к движению, к изменению поведения для того, чтобы восстановить изменившиеся параметры внутренней среды организма. Под влиянием мотивации и при участии коры головного мозга происходит формирование программы конкретного поведения, в которую включается и изменение дыхания.
Особенно велика роль дыхания в терморегуляции, потому что при изменении паттерна дыхания возможно или увеличить, или уменьшить теплоотдачу. При повышении температуры повышается частота дыхания, за счет этого происходит увеличение испарения воды и некоторое охлаждение организма. Умеренное охлаждение тоже приводит к увеличению частоты дыхания. Это связано с возбуждением задних областей гипоталамуса и общим повышением активности и возбудимости структур мозга, в том числе и дыхательного центра. При значительном охлаждении угнетается активность нейронов дыхательного центра, глубина и частота дыхания уменьшаются, что позволяет уменьшить потери тепла.
Из изложенного становится ясно, что в дыхательный центр, наряду с информацией от хеморецепторов и от рецепторов растяжения легких, поступает поток информации обо всем, что происходит в организме и окружающей среде. Разница заключается в том, что афферентные сигналы от разнообразных рецептивных полей и от анализаторов поступают не непосредственно в дыхательный центр, а в различные отделы головного мозга. Эти отделы, в свою очередь, оказывают возбуждающее или тормозное влияние и на дыхательный центр, и на другие функциональные системы (например, система кровообращения). В различных ситуациях центры головного мозга образуют с дыхательным центром временные функционально подвижные ассоциации (функциональные системы по А.П. Анохину), которые обеспечивают полноценное регулирование дыхания в соответствии с потребностями организма.
Кора больших полушарий
Роль коры больших полушарий в регуляции дыхания изучена в экспериментах на животных с электрическим раздражением различных зон коры и с их удалением. Оказалось, что у бескорковых животных с нормальным дыхательным ритмом в покое наступает резко выраженная и длительная одышка уже после нескольких шагов. Следовательно, для приспособления дыхания к мышечной деятельности требуется участие высших отделов центральной нервной системы. В коре мозга нет определенных участков, четко изменяющих деятельность дыхательного центра. Раздражение большинства участков коры приводит к изменению паттерна дыхания. Вместе с тем, наиболее существенные изменения дыхания наблюдаются при раздражении соматосенсорной области коры. Это и понятно, именно мышечная деятельность требует наиболее существенного изменения дыхания. Во время работы мышцам необходимо очень большое количество кислорода. Если в покое потребление кислорода составляет 250-300 мл/минуту, то при быстрой ходьбе оно возрастает до 2.5 литров, а при тяжелой мышечной работе до 4л/минуту. Обеспечение мышц кислородом достигается совместной деятельностью систем кровообращения и дыхательной.
Механизмы регуляции вентиляции при мышечной работе сложны. Казалось бы, увеличение МОД можно объяснить повышением частоты импульсов с хеморецепторов продолговатого мозга и каротидных синусов, которое возникает в результате повышения напряжения углекислого газа и снижения напряжения кислорода в крови. Однако вентиляция легких всегда увеличивается в начале работы, когда газовый состав крови еще не успел измениться. Следовательно, гиперпноэ возникает под влиянием нервных факторов. Кора больших полушарий, вызывая произвольные движения, активирует и деятельность дыхательного центра и непосредственно, и через гипоталамус. В дополнение к этой стимуляции возбуждение дыхательного центра увеличивается под влиянием импульсов от проприорецепторов работающих мышц. Эти импульсы возникают даже в тех случаях, когда работа мышц осуществляется пассивно, без затрат кислорода и выделения углекислого газа (например, экспериментатор сгибает конечности испытуемого).
Позднее, во время продолжающейся физической работы, происходит более медленное увеличение вентиляции легких. Это увеличение вентиляции уже обусловлено раздражением артериальных и центральных хеморецепторов. Однако и здесь не все так просто. Выяснилось, что даже при физической работе высокой интенсивности напряжение кислорода и углекислого газа в крови может не измениться (а углекислого газа даже снизиться в результате возросшей вентиляции легких). Даже удаление каротидных телец не устраняет увеличения легочной вентиляции во время физической работы, и тем не менее, сигналы от хеморецепторов имеют существенное значение в увеличении МОД во время физической работы. Оказывается, что во время работы увеличивается чувствительность дыхательного центра к гиперкапнии и гипоксии, возрастает и возбудимость хеморецепторов. Имеет значение и повышение температуры тела: этот фактор через центры гипоталамуса увеличивает частоту дыхания.
Кортикальные влияния на дыхание отчетливо проявляются при тренировке к выполнению одной и той же работы. Постепенно колебания дыхания становятся меньше, дыхание делается более ровным. Если много раз выполняется работа в одинаковом темпе, но с различной интенсивностью, то изменения вентиляции при переходе на новый уровень происходят быстрее, вырабатывается динамический стереотип, в котором вентиляция легких имеет волнообразный характер. Из этого примера видно, что важные приспособительные изменения дыхания осуществляются посредством выработки условных рефлексов. Примером такого условно-рефлекторного изменения ритма дыхания может быть стартовый рефлекс у спортсменов.
Особенно отчетливо роль коры головного мозга проявляется в произвольном управлении дыханием. Своеобразие функции внешнего дыхания заключается в том, что она одновременно и обладает автоматизмом, и произвольно управляема. Человек прекрасно дышит и во сне, и под наркозом, однако любой человек может произвольно остановить дыхание, изменить его частоту и глубину. Произвольное управление дыханием возможно потому, что в коре есть представительство дыхательных мышц и корковомедуллярные нисходящие влияния на мотонейроны дыхательных мышц. Возможность произвольного управления дыханием ограничена определенными пределами изменений напряжения кислорода и углекислого газа, а так же рН крови. При чрезмерной задержке дыхания возникает стимул, который возвращает дыхание под контроль дыхательного центра. Значение возможности произвольного контроля дыхания для человека трудно переоценить - именно благодаря такой возможности человек приобрел одну из немногих, отличающих его от животных, функций - способность к членораздельной речи.
Заключение
Дыхательный центр получает большой объем информации и о газовом составе крови, и о состоянии дыхательной системы. В соответствии с этой информацией изменяется легочная вентиляция. Вместе с тем дыхательный центр способен обеспечивать смену дыхательных фаз за счет своих собственных механизмов возбуждения нейронов, т.е. обладает определенным автоматией. Автоматия дыхательного центра отличается рядом признаков от автоматии сердечного водителя ритма.
Периодическое возбуждение дыхательного центра обусловлено взаимодействием большого числа нервных клеток, объединенных в нейронные сети. Среди них особую роль играют тормозные нейроны. В дыхательном центре есть несколько популяций инспираторных и экспираторных нейронов соответственно фазам дыхательного цикла. Среди нейронов есть проприобульбарные нейроны, которые организуют паттерн дыхания, определяя уровнем своего возбуждения длительность фаз дыхательного цикла. Бульбоспинальные нейроны являются преимущественно премоторными по отношению к мотонейронам диафрагмального нерва и нервам межреберных мышц. Смена дыхательных фаз осуществляется путем постепенного возбуждения одних групп нейронов и торможения других. Для осуществления смены дыхательных фаз необходимо влияние пневмотаксического центра, расположенного в районе варолиевого моста среднего мозга.
Для автоматической деятельности дыхательного центра необходимо постоянное (тоническое) поступление к нему сигналов, повышающих возбудимость дыхательных нейронов. Сигналы, поступающие от хеморецепторов продолговатого мозга и каротидного синуса, а так же от рецепторов растяжения легких изменяют активность дыхательных нейронов в соответствии с газовым составом крови и состояние дыхательной системы. Информация от этих рецепторов лежит в основе регуляции по принципу отклонения. Афферентные сигналы, поступающие от всех рецептивных полей через ретикулярную формацию, гипоталамус и кору головного мозга изменяют активность дыхательных нейронов в соответствии с состоянием внутренней среды организма и окружающей среды, поведением. Таким образом, осуществляется регуляция дыхания по принципу возмущения.
Автоматическая деятельность дыхательного центра человека находится под значительным произвольным контролем.Человек может в широких пределах произвольно изменять частоту и глубину дыхания. Вместе с тем, этот произвольный контроль ограничен влияниями от центральных и периферических хеморецепторов, рецепторов растяжения легких, ирритантных и других механорецепторов.
Объединим на рисунке 25 основные структуры, принимающие участие в регуляции дыхания и по принципу отклонения, и по принципу возмущения, и суммируем их роль в регуляции дыхания.
Рисунок 25. Отделы головного мозга,
Перерезка на уровне выше варолиева моста не изменяет дыхания покоя, но резко искажает адаптивные реакции дыхательной системы.
Перерезка на уровне между верхней и средней частью моста, приводит к изменению дыхания по типу айпнезиса,состоящее в продолжительном вдохе и резком выдохе, что связано с устранением тормозного влияния верхней части моста на инспираторные нейроны.
Перерезка на уровне между мостом и продолговатым мозгом приводит к изменению дыхания по типу гаспинга. Ритм дыхания становится неправильным. Нарушается соотношение продолжительности вдоха и выдоха. Кратковременный вдох и продолжительный выдох.
Перерезка между грудным и шейным отделами спинного мозга сохраняет ритм дыхания, но снижает мощность как вдоха, так и выдоха.
Исходя из результатов опытов с перерезкой центральной нервной системы, можно определить роль разных отделов ЦНС в регуляции внешнего дыхания.
Спинной мозг - обеспечивает эфферентную иннервацию дыхательных мышц (III - IV сегменты шейного отдела иннервируют диафрагму, грудные сегменты - межреберные), является исполнительной структурой, не обладающей самостоятельной активностью.
Продолговатый мозг - является генератором центрального дыхательного ритма.
Варолиев мост - обеспечивает правильность центрального дыхательного ритма, оптимальное соотношение между продолжительностью вдоха и выдоха. В нем расположены пневмотаксический и айпнестический центры.
Гипоталамус - осуществляет безусловнорефлекторные адаптивные реакции внешнего дыхания.
Высшие отделы мозга (новая кора и лимбическая система) - обеспечивают условнорефлекторные механизмы адаптивных реакций дыхания.
3. Механизм центрального дыхательного ритма.Центральный дыхательный ритм генерируется бульбарным отделом дыхательного центра. Для него свойственна автоматия, которая проявляется в периодической смене возбуждения и торможения инстираторных и экспираторных нейронов. Доказательством автоматии является существование слабых дыхательных движений у плода на последних месяцах беременности, электрофизиологическая регистрация смены возбуждения и торможения нейронов дыхательного центра.
Дыхательные нейроны локализованы в парных вентральных и дорсальных ядрах дна 4 желудочка продолговатого мозга.
Дорсальное ядро содержит преимущественно инспираторные нейроны. Их аксоны образуют синапсы на мотонейронах, иннервирующих мышцы диафрагмы. Благодаря преимущественному расположению инспираторных нейронов в дорсальных ядрах их условно называют центром вдоха (инспираторным).
Вентральные ядра содержат и инспираторные, и экспираторные нейроны. Их аксоны образуют синаптические контакты на мотонейронах межреберных мышц и мышц брюшной стенки. Обоюдное ядро связано с вентральными ядрами, обеспечивает согласование сокращения мышц глотки и гортани с дыхательным циклом. Между инспираторными и экспираторными нейронами существуют антагонистические отношения, которые проявляются в том, что возбуждение одних приводит к торможению других.
Одна из гипотез происхождения центрального дыхательного ритма (периодической смены возбуждения инспираторных и экспираторных нейронов) предполагает неоднородность популяции инспираторных нейронов - Ra и Rb.
Ra нейроны активны только на вдохе, а Rb нейроны активны и на вдохе, и на выдохе, причем их активность возрастает к концу вдоха, усиливается при увеличении афферентной импульсации от механорецепторов растяжения легких. Предположено, что Rb группа нейронов тормозит активность Ra нейронов.
Согласно этой гипотезе, упрощенный механизм центрального дыхательного ритма реализуется следующим образом. Предполагается, что Ra нейроны обладают спонтанной активностью. Их возбуждение через синаптические контакты вызывает возбуждение мотонейронов инспираторных мышц - происходит вдох. Посредством коллатералей возбуждение передается на Rb нейроны, которые посредством вставочных тормозных нейронов тормозят активность Ra нейронов. Они переходят в состояние торможения, перестают возбуждать инспираторные мотонейроны и происходит выдох. Торможение Ra нейронов вызывает прекращение активации Rb нейронов, которые в свою очередь перестают тормозить спонтанно активные Ra нейроны. Они возбуждаются, и происходит следующий вдох.
Центральный дыхательный ритм определяется активностью бульбарных нейронов, модулируется другими отделами ЦНС и афферентной импульсацией с периферии.
4. Значение центральных и периферических хеморецепторов. Частота и глубина дыхания определяются тонусом дыхательного центра, который зависит от общего состояния центральной нервной системы. Экспериментально доказано изменение активности дыхательного центра при сдвигах в газовом составе крови. Первые экспериментальные данные, доказывающие гуморальную регуляцию дыхания, были получены в опытах с перекрестным кровообращением (Фредерик, 1890 г). Изменение афферентной импульсации от хеморецепторов вызывает адекватное изменение частоты и глубины дыхания. Тем самым сохраняется постоянство газового состава артериальной крови при значительных колебаниях парциального давления кислорода в атмосферном воздухе (от 80 до 300 мм.рт.ст.).
· Нормоксия - нормальное напряжение кислорода в крови.
· Гипоксемия - пониженное напряжение кислорода в крови.
· Гипероксия - повышенное содержание кислорода в крови.
· Гипоксия - сниженное содержание кислорода в организме.
· Нормокапния - нормальное содержание в крови СО2..
· Гиперкапния - повышенное содержаение в крови СО2..
· Гипокапния - пониженное содержание в крови СО2..
Типы вентиляции легких во многом зависят от напряжения газов.
· Нормовентиляция или эйпноэ (РСО2 в альвеолах и артериальной крови поддерживается на уровне 40 мм рт.ст.).
· Гипервентиляция (РСО2 больше 40 мм.ртст).
· Гиповентиляция (РСО2 меньше 40 мм.рт.ст.).
· Повышенная вентиляция (увеличение минутного объема легких независимо от напряжения углекислого газа и кислорода, например, при мышечной работе).
· Эупноэ - вентиляция с ощущением комфорта.
· Тахипноэ - увеличение частоты дыхания.
· Брадипноэ - уменьшение частоты дыхания.
· Гиперпноэ - увеличение глубины дыхания.
· Апноэ - остановка дыхания, может возникать при гипоксии вследствие снижения оксигенации головного мозга.
· Ортопноэ - одышка в связи с застоем крови в малом круге кровообращения.
· Асфиксия - остановка дыхания в связи с параличом дыхательного центра.
Влияние РСО2 . При повышении напряжения углекислого газа в крови с 40 до 60 мм рт.ст. вентиляция легких возрастает с 6-7 л/мин до 70 л/мин. При дальнейшем увеличении напряжения газа минутный объем вентиляции не возрастает.
Влияние рН. Уменьшение рН крови на 0,1 увеличивает вентиляцию легких на 2л/мин (при РСО2 40 мм рт.ст.). Газовый ацидоз вызывает более значимое увеличение легочной вентиляции, чем метаболический.
Влияние РО2. Гипоксемия приводит к увеличению легочной вентиляции, но в гораздо меньшей степени, чем гиперкапния.
В условиях физиологической нормы регуляция внешнего дыхания обеспечивается изменениями рН и РСО2 в крови, но не РО2.
При патологии (отравлении барбитуратами), когда возбудимость дыхательного центра и его чувствительность к РСО2 уменьшается, артериальная гипоксемия становится основным стимулятором деятельности дыхательного центра. В этой ситуации назначение больным кислорода может стать опасным для жизни, так как может вызвать остановку дыхания.
Информацию о газовом составе крови дыхательный центр получает посредством периферических и центральных хеморецепторов.
Периферические хеморецепторы. Расположены в важнейших сосудистых рефлексогенных зонах - дуге аорты (аортальное тельце) и каротидном синусе. Основными являются рецепторы каротидного синуса. Реагируют на гиперкапнию, ацидоз и гипоксемию. При гипероксии их чувствительность к рН и РСО2 снижается. Преимущественно поставляют информацию в дыхательный центр о РО2. Афферентная импульсация от периферических хеморецепторов преимущественно усиливает активность инспираторных нейронов.
Центральные хеморецепторы. Находятся в продолговатом мозге. Реагируют на снижение рН ликвора. Их чувствительность очень высокая. Снижение рН на 0,01 обеспечивает увеличение легочной вентиляции на 4 л/мин. При снижении рН увеличивается общая интенсивность центрального дыхательного ритма (тонус дыхательного центра). Раздражение центральных хеморецепторов приводит к активации как инспираторных, так и экспираторных нейронов.
В состоянии физиологической нормы основным механизмом регуляции деятельности дыхательного центра является изменение рН ликвора.
5. Механорецепторы легких в регуляции дыхания.В легких выделяют три группы механорецепторов: рецепторы растяжения, ирритантные рецепторы, юкстаальвеолярные/капиллярные/ рецепторы.
Рецепторы растяжения. Находятся в гладкомышечном слое воздухоносных путей, отсутствуют в альвеолах и плевре. Обладают низкой способностью к адаптации. Афферентная импульсация от них увеличивается при растяжении легких на вдохе. Их возбудимость снижается при снижении РСО2 в альвеолярном воздухе.
Возбуждение от этой группы рецепторов передается по чувствительным волокнам вагуса в продолговатый мозг, усиливает экспираторную активность дыхательного центра и снижает инспираторную (активирует Rb нейроны и тормозит Raнейроны).
Перерезка вагуса приводит к снижению частоты, но увеличению глубины дыхания.
Рефлексы Геринга-Брейера ( 1868).
– Инспираторно тормозной рефлекс. Состоит в торможении входа при искусственном раздувании легких на вдохе.
– Экспираторно облегчающий рефлекс. Состоит в задержке выдоха при раздувании легких на выдохе.
– Парадоксальный эффект Хеда. Проявляется в судорожном вдохе при быстром раздувании легких.
– Рефлекс на спадение легких. Значительное уменьшение объема легких приводит к увеличению инспираторной активности.
Рецепторы растяжения являются важной составляющей в механизме ограничения продолжительности вдоха.
– Ирритантные рецепторы. Расположены в эпителии и субэпителии воздухоносных путей. Обладают высокой способностью к адаптации. Являются механо- и хемочувствительными. Раздражаются при растяжении легких, частицами пыли, парами аммиака, табачным дымом, едкими веществами. При их раздражении осуществляются защитные дыхательные рефлексы (кашель, чихание).
– Юкстаальвеолярные/капиллярные/ рецепторы. Находятся в капиллярах малого круга кровообращения. Активируются при гипертензии малого круга, действии гистамина, никотина, отеке легких и повреждении легочной ткани. Афферентная импульсация поступает в продолговатый мозг, вызывает изменение дыхания по типу ортопноэ. Одновременно происходит рефлекторная бронхоконстрикция.
6. Сопряженные рефлексы дыхания. Осуществляются при раздражении других экстеро- и интерорецепторов.
Артериальные прессорецепторы. Их раздражение увеличением давления в артериях приводит к слабому снижению вентиляции легких. Падение давления приводит к обратному эффекту.
Пропреорецепторы дыхательных мышц. Их количество на единицу объема мышечной ткани больше чем в большинстве скелетных мышц. Афферентация от инспираторных мышц возрастает при вдохе и снижается при выдохе. Является вспомогательным фактором для ограничения продолжительности вдоха.
Рецепторы верхних дыхательных путей. Сильное раздражение слизистой полости носа водой и едкими веществами приводит к временной остановке дыхания (рефлекс ныряльщиков). Важное значение имеют в акте глотания и защитных дыхательных рефлексах.
Влияние моторных зон ЦНС. Является определяющим при максимальной вентиляции легких в условиях физической нагрузки, когда МОД ( минутный объем дыхания ) возрастает до 120 л/мин. Эффект связан с активацией гипоталамических механизмов интеграции моторной активности и её вегетативного обеспечения.
Экстеротерморецепторы. Сильное возбуждение вызывает увеличение МОД ( минутного объема дыхания ).
Интеротерморецепторы. Повышение и незначительное понижение температуры тела приводит к усилению дыхания. Гипотермия вызывает снижение МОД.
Болевые экстерорецепторы. Их раздражение усиливает дыхание.
Гормональные влияния. Адреналин увеличивает МОД при стрессорных реакциях. Прогестерон способствует усилению дыхания при беременности.
7. Механизм первого вдоха новорожденного.К моменту рождения у ребенка сформированы структуры дыхательного центра, существует центральный дыхательный ритм, который проявляется во внутриутробных дыхательных движениях плода. Тонус нервной системы и дыхательного центра низкий. Оксигенация крови и выведение углекислого газа целиком определяется организмом матери. В процессе родовой деятельности прерывается связь ребенка с материнским организмом. Развивающаяся гиперкапния, гипоксемия и функциональный ацидоз посредством центральных и периферических хеморецепторов вызывают увеличение тонуса дыхательного центра. Большое значение в усилении инспираторной активности играет сильное раздражение тактильных, температурных и болевых экстероцецепторов. Сильное возбуждение инспираторных нейронов активирует инспираторные мотонейроны. Происходит вдох. В связи с существованием и активацией центрального дыхательного ритма и резким увеличением афферентации от механорецепторов растяжения возбуждение инспираторных нейронов уменьшается, но возбуждаются экспираторные нейроны. Возникает первый активный выдох. В связи с автоматией дыхательного центра вслед за ним происходят последующие вдох и выдох. Через несколько дыхательных экскурсий газовый состав крови нормализуется.
Читайте также:
