
Торможение в цнс механизм пресинаптического торможения
Лекция 6. Распространение возбуждения в ЦНС. Процессы торможения в ЦНС
Распространение возбуждения в ЦНС
Особенности распространения возбуждения в ЦНС объясняются ее нейронным строением:
· Наличие химических синапсов,
· Многократное ветвление аксонов,
· Наличие замкнутых нейронных сетей.
Как следствие, распространение возбуждения в ЦНС имеет следующие характеристики:
Одностороннее распространение возбуждения в нейронных цепях, рефлекторных дугах (синапсы).
2. Замедленное распространение возбуждения в ЦНС(в сравнении с нервным волокном). Множество хим. синапсов, в каждом до возникновения ВПСП (возбуждающий постсинаптический потенциал) синаптическая задержка 0,5 -2 мс.
3. Иррадиация (дивергенция=расхождение) возбуждения в ЦНС.Из-за ветвления аксонов, они могут устанавливать многочисленные связи с другими нейронами, кроме того, есть вставочные нейроны, которые также ветвятся.
Пример: Спинальная лягушка (у которой нарушена связь спинного мозга с головным). Слабое раздражение вызывает сгибание одной конечности, сильное – всех.
Значение: дивергенция расширяет сферу действия одного нейрона. Один нейрон коры больших полушарий может участвовать в возбуждении 5000 нейронов, в других отделах ЦНС – 1000.
4. Конвергенция (=схождение) возбуждения (=принцип общего конечного пути=воронка Шеррингтона). Возбуждение к одному и тому же нейрону может приходить по нескольким путям, т.к.
А) у нейронов множество аксонных ответвлений (коллатералей),
Б) в ЦНС множество вставочных нейронов,
В) в организме афферентных входов в несколько раз больше, чем эфферентных выходов.
Именно конвергенция обеспечивает пространственную суммацию многих ВПСП для возникновения ПД (или же блокирование – при преобладании тормозных влияний).
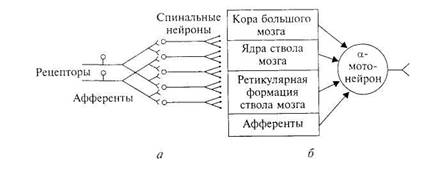
Пример: к одному спинальному α-мотонейрону подходят первичные афференты, нисходящие пути многих вышележащих центров ЦНС и т.д.
Значение:один мотонейрон может участвовать в нескольких реакциях: например, участие мотонейрона, иннервирующего мышцы глотки, в глотании, кашле, чихании.
5. 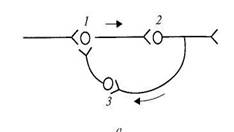
Циркуляция возбуждения по замкнутым нейронным цепям.Может длиться минуты, часы. Обеспечивает последействие.Наиболее вероятный механизм кратковременной памяти.
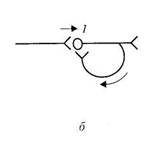
Рис. Цепь возбуждающих нейронов (Лоренто де-Но) Рис. Один нейрон (И.С. Беритов)
6. Распространение возбуждения в ЦНС легко блокируется фармакологическими препаратами (блокаторы ионных каналов, блокаторы рецепторов постсинаптической мембраны).
Процесс торможения в ЦНС
Для нормальной деятельности ЦНС необходимо строго упорядоченное чередование активности определённых групп нейронов, соединённых друг с другом топографически точными (как на карте) связями. Предупреждение распространения возбуждения на соседние, не участвующие в процессе, нейроны обеспечивается тормозными нейронами.
Торможение– это активный нервный процесс, результатом которого является прекращение или ослабление возбуждения.
Пример роли торможения: Если экспериментальному животному ввести некоторое количество стрихнина (это алкалоид семени чилибухи или рвотного ореха), блокирующего только одну разновидность тормозных синапсов в центральной нервной системе, то начнётся неограниченное распространение возбуждения в ответ на любой раздражитель, что приведёт к неупорядоченной активности нейронов, затем возникнут мышечные судороги, конвульсии и, наконец, смерть.
Торможение всегда является местным процессом, оно не может, подобно возбуждению, распространяться от одной клетки к другой. Торможение лишь угнетает процесс возбуждения или препятствует самому возникновению возбуждения.
Тормозные нейроны есть во всех областях мозга, например, в спинном мозге – тормозные клетки Реншоу, в коре мозжечка – нейроны Пуркинье, звёздчатые клетки и т.д.
Нисходящее (центральное) торможение впервые описал И. М. Сеченов: он обнаружил, что рефлексы спинного мозга у лягушки замедляются, если её промежуточный мозг раздражать кристалликом поваренной соли.
Так, кора больших полушарий оказывает постоянные тонические тормозные влияния на нижележащие подкорковые структуры (При аффекте подкорка выходит из-под этих влияний).
У млекопитающих в свою очередь торможение коры больших полушарий (сонное состояние) вызывает ретикулярная формация среднего мозга.
Раздражение хвостатого ядра (часть полосатого тела – Базальные ядра) вызывает общее торможение животного, прекращение начатых условных рефлексов и т.д.
Пресинаптическое и постсинаптическое торможение
Торможение, если рассматривать его на микроуровне, разделяется на пресинаптическое и постсинаптическое.
Пресинаптическое торможение
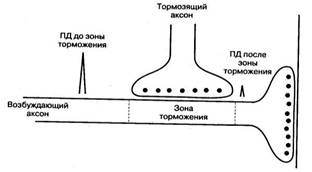
Наблюдается в аксо-аксональных синапсах: здесь аксон тормозного нейрона образует синапс на окончании возбуждающего нейрона. В таких синапсах обычно используются каналы для ионов хлора – их вход в окончание возбуждающего нейрона уменьшает амплитуду проводящихся по нему потенциалов действия. В связи с этим уменьшается количество медиатора, который выделяется в окончании аксона и, соответственно величина ВПСП.
Пресинаптическое торможение является избирательным – оно блокирует лишь один возбуждающий вход и постсинаптический нейрон сохраняет возможность возбуждаться другими, не подвергнутыми торможению нейронами.
Важно: Наряду с пресинаптическим торможением встречается и пресинаптическое усиление, когда в аксо-аксональном синапсе действует медиатор, повышающий эффективность возбуждающего нейрона. Т.е. аксо-аксональные синапсы являются модуляторами: не имея прямого влияния на зону возникновения ПД, они изменяют возбудимость ЦНС.
Постсинаптическое торможение
Постсинаптическое торможение происходит в тормозных синапсах. Как рассматривалось ранее, в них также выделяется медиатор и открываются хемозависимые каналы. Но каналы предназначаются не для натрия, как в возбуждающих синапсах, а для прохождения ионов хлора (внутрь) или ионов калия (наружу). Происходит гиперполяризация мембраны. Из состояния гиперполяризации возбудить нейрон труднее.
Возникающий гиперполяризующий сдвиг и называется тормозным постсинаптическим потенциалом (ТПСП).
Виды постсинаптического торможения:
· латеральное и т.д.
Возвратное торможение позволяет регулировать выходную активность нейрона по принципу отрицательной обратной связи. Здесь возбуждающий какую-либо клетку нейрон одной из коллатералей своего аксона действует ещё и на вставочный тормозной нейрон, который начинает тормозить активность самой возбуждающей клетки.
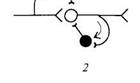
(здесь под цифрой 2 – возвратное, заштрихованные нейроны – тормозные, а светлые – возбуждающие)
Так, например, мотонейрон спинного мозга возбуждает мышечные волокна, а другая коллатераль его аксона возбуждает клетку Реншоу, которая тормозит активность самого мотонейрона
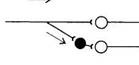
Благодаря реципрокному торможению мышцы-антагонисты не могут сокращаться одновременно и, если для совершения движения сокращаются сгибатели, то разгибатели должны расслабляться.

Латеральное торможение. Тормозные вставочные нейроны активируются импульсами из возбужденного центра и влияют на соседние (расположенные сбоку) клетки с такими же функциями. В этих соседних клетках развивается очень глубокое торможение. Так образуется тормозная зона вокруг возбужденных нейронов.
3. Роль торможения в деятельности нервной системы:
1. Оба вида торможения во всех их разновидностях выполняют охранительную роль. Иначе происходит истощение медиаторов, прекращение или нарушение работы ЦНС.
2. Торможение играет важную роль в обработке поступающей в ЦНС информации (особо – пресинаптическое торможение, которое сдерживает количество приходящих к нейрону импульсов). В случае латерального торможения – позволяет выделить существенные сигналы из фона.
3. Торможение – важный фактор обеспечения координационной деятельности ЦНС.
Дата добавления: 2018-04-05 ; просмотров: 3031 ;
Тема: Торможение в ЦНС. Принципы координационной деятельности ЦНС.
Учебные и воспитательные цели:
Изучить природу, функциональное значение, структурно-функциональную организацию торможения в центральной нервной системе.
- Физиология человека. Под ред. В.М.Покровского, Г.Ф.Коротько.М., Медицина. – 2000.-Т..1 - С 115 – 126.
Мультимедийная презентация 34 слайда.
РАСЧЕТ УЧЕБНОГО ВРЕМЕНИ
| № п/п | Перечень учебных вопросов | Количество выделяемого времени в минутах |
| 1. | Введение | |
| Торможение в ЦНС. Природа торможения. Виды торможения. | ||
| Первичное постсинаптическое торможение | ||
| Пресинаптическое торможение | ||
| Вторичное торможение | ||
| Механизмы координации рефлекторной деятельности | ||
| Компенсация функций и пластичность нервных центров | ||
| Заключение |
Торможение в ЦНС. Природа торможения. Виды торможения. Взаимосвязь возбуждения и торможения в ЦНС.
Торможение в ЦНС - особый нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения.
Классификация и механизмы центрального торможения:
1. Первичное постсинаптическое торможение - торможение , не- связанное с первоначальным процессом возбуждения и развивающееся в результате активации специальных тормозных структур. Тормозные синапсы образуют в своих окончаниях тормозной медиатор (ГАМК, глицин, в отдельных синапсах ЦНС роль тормозного медиатора может играть ацетилхолин). На постсинаптической мембране развивается тормозной постсинаптический потенциал (ТПСП), снижающий возбудимость мембраны постсинаптического нейрона. Тормозными нейронами могут служить только вставочные нейроны, афферентные нейроны всегда являются возбуждающими. В зависимости от вида тормозных нейронов и структурной организации нейронной сети постсинаптическое торможение подразделяется на:
Реципрокное торможение. Оно лежит в основе функционирования мышц-антагонистов и обеспечивает расслабление мышцы в момент сокращения мышцы-антагониста. Афферентное волокно, проводящее возбуждение от проприорецепторов мышц (например, сгибателей), в спинном мозге делится на две ветви: одна из них образует синапс на мотонейроне, иннервирующем мышцу-сгибатель, а другая - на вставочном, тормозном, образующем тормозной синапс на мотонейроне, иннервирующем мышцу-разгибатель. В результате возбуждение, приходящее по афферентному волокну, вызывает возбуждение мотонейрона, иннервирующего сгибатель и торможение мотонейрона мышцы-разгибателя.
Возвратное торможение. Оно реализуется через тормозные клетки Реншоу, открытые в спинном мозге. Аксоны мотонейронов передних рогов отдают коллатераль на тормозной нейрон Реншоу, аксоны которого возвращаются на тот же мотонейрон, образуя на нем тормозные синапсы. Таким образом формируется контур с отрицательной обратной связью, позволяющий стабилизировать частоту разрядов мотонейрона.
Центральное (Сеченовское) торможение. Оно осуществляется тормозными вставочными нейронами, через которые реализуется влияние на мотонейрон спинного мозга возбуждения, возникающего в зрительных буграх под влиянием их раздражения. На мотонейроне спинного мозга суммируются ВПСП, возникающие в болевых рецепторах конечности и ТПСП, возникающие в тормозных нейронах под влиянием возбуждения таламуса и ретикулярной формации. В результате время защитного сгибательного рефлекса возрастает.
Латеральное торможение осуществляется с помощью тормозных вставочных нейронов в параллельных нейронных сетях.
Первичное пресинаптическое торможение развивается в терминальных отделах аксонов (перед пресинаптической структурой) под влиянием специальных аксо-аксональных тормозных синапсов. Медиатор этих синапсов вызывает деполяризацию мембраны терминалей и приводит их в состояние, подобное катодической депрессии Вериго. Мембрана в области такого бокового синапса препятствует проведению потенциалов действия к пресинаптической мембране, активность синапса уменьшается.
Пресинаптическое торможение
Пресинаптическим торможением называют снижение или выключение активности клетки за счет синаптического торможения оканчивающейся на ней возбуждающей терминали. Явление пресинаптического торможения зафиксировали Гассер и Грэхем в 1933 г., наблюдая эффект развития торможения сгибательных рефлексов при раздражении других корешков. Данный вид торможения термином “пресинаптическое торможение” впервые обозначили Фрэнк и Фуортес в 1957 г.

Рис. 1. Электронная микрофотография пресинаптических тормозных синапсов. Обозначения: пре. и пост. – пре- и постсинаптический контакт нейрона.
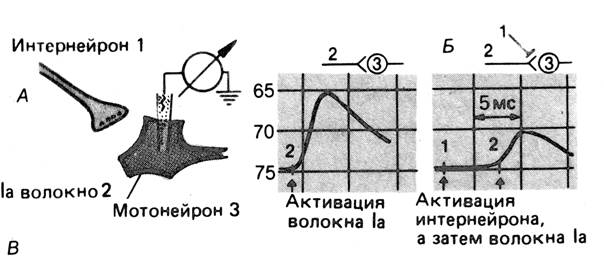
Рис. 2. Пресинаптическое торможение.
А. Схема эксперимента для демонстрации пресинаптического торможения моносинаптических ВПСП в мотонейроне.
Б. ВПСП при стимуляции гомонимных волокон Ia без (слева) и после (справа) предварительной активации пресинаптических тормозных интернейронов.
По “Физиология человека” под ред Р.Шмидта. – 1996. – Т.1. – С.59.
Стимуляция тормозного интернейрона подавляет моносинап-тический ВПСП мотонейрона.

Рис. 3. Подавление моносинаптического ВПСП при синаптическом торможении.
По Дж. Экклс. Физиология синапсов. – М.: Мир. – 1966. – С279.
Развитие этого подавления ВПСП протекает с латентным периодом, равным 5 мс, максимум достигается через 20 мс, а общая длительность превосходит 200 мс.

Рис. 4. Временной ход пресинаптического торможения моносинаптических возбуждающих ВПСП.
По “Физиология человека” под ред Р.Шмидта. – 1996. – Т.1. – С.59.
Увеличение частоты предварительных раздражений изменяет характер подавления. В частности, одна серия стимуляции с частотой 200-300 импульсов в секунду вызывает максимальное подавление менее чем на 10%, а две серии – подавление менее чем на 20%. При пресинаптическом торможении подавление моносинаптического ВПСП не связано с какими-либо изменениями их временных параметров.
Тормозные синапсы на окончаниях волокон Ia обеспечивают довольно значительную деполяризацию, называемую деполяризацией первичных афферентов, или же первичную эфферентную деполяризацию(ПАД). В спинном мозгу ПАД обнаруживает длительную фазу (до 25 мс) нарастания до закругленной вершины и характеризуется большей продолжительностью по сравнению с постсинаптическими процессами. Большая продолжительнсоть ПАД объясняется или длительным действием медиатора, или медленным, пассивным снижением деполяризации вследствие большой электрической постоянной времени мембраны. Пассивно снижающаяся компонента ПАД снимается импульсом, распространяющимся по афферентному волокну до его центральных окончаний.
Существует соответствие во всех отношениях между наблюдаемой деполяризацией первичных афферентных волокон и подавлением их синаптического возбуждающего действия.
Пресинаптическая деполяризация афферентов уменьшает величину их пресинаптического спайкового потенциала и таким образом уменьшает вызываемый им ВПСП. По данным Каца (1962), снижение спайкового потенциала на 5 мВ приводит к снижению выброса квантов медиатора и к снижению ВПСП до 50% и менее.
Характер ПАД в различных нейронах отличается по своим характеристикам. В целом временные параметры сравнимы. ПАД волокон кожного нерва отличается большей величиной амплитуды на одиночные раздражения с более коротким латентным периодом (около 2 мс), максимум также достигается ранее, чем в случае ПАД, вызываемых ритмическим раздражением нервных волокон, идущих от мышц. ПАД в клиновидном ядре имеет короткий латентный период (около 2 мс) и быстрый подъем до максимума.
Тормозные синапсы имеют химическую природу, медиатором в них служит ГАМК. Деполяризация первичных афферентов инактивирует возбуждающие натриевые каналы.Шунтирование натриевых каналов снижает амплитуду пресинаптического потенциала действия. В результате синаптическая передача моторного импульса ослабляется или исключается.
Во всех типах возбуждающих синапсов обнаруживается тесная зависимость между деполяризацией пресинаптических волокон и торможением синаптической передачи. Пресинаптическое торможение оказывает влияние на возбуждающие синапсы всех типов, образуемые афферентными волокнами, входящими в спинной мозг, групп Ia, Ib и II, отходящими от мышц, и альфа-волокнами кожного нерва. Это торможение влияет не только на местные спинномозговые рефлексы, но также и на синаптические передачи в восходящих путях как от кожных афферентов, так и на спиноцеребеллярные. Кроме того, пресинаптическое торможение влияет на синаптические передачи задних столбов в ядра нежного и клиновидного пучков. Нисходящие импульсы из коры головного мозга и ствола мозга также оказывают пресинаптическое тормозное влияние на волокна группы I и кожные афферентные волокна в спинном мозгу и клиновидном ядре. Обнаружено пресинаптическое торможение вторичных афферентных волокон отходящих от клиновидного ядра и имеющих переключение в таламусе. Синапсы с пресинаптическим торможением обнаружены в связанном с таламусом ядре мозга – латеральном коленчатом теле. В коре головного мозга не выявлено синаптических структур, которые могли бы осуществлять пресинаптическое торможение. На этих высших уровнях нервной системы доминирует постсинаптическое торможение. Пресинаптическое торможение действует как отрицательная обратная связь, уменьшая приток сенсорной информации в центральную нервную систему. Обычно эта отрицательная обратная связь не имеет точной топографии, но обычно концентрируется в пределах одной сенсорной модальности.
Пресинаптическое торможение служит механизмом регуляции двигательных систем спинного мозга. Его особенностью является возможность специфического воздействия на отдельные синаптические входы без изменений возбудимости всей клетки. Таким образом, избыточная информация устраняется еще до того, как достигает места интеграции клеточного тела нейрона.
2. Вторичное торможение не связано с тормозными структурами, является следствием предшествующего возбуждения. Пессимальноеторможение (открыто Н.Е.Введенским в 1886 г.) развивается в полисинаптических рефлекторных дугах при чрезмерной активации центральных нейронов и играет предохранительную роль. Оно выражается в стойкой деполяризации мембраны, приводящей к инактивации натриевых каналов. "Торможение вслед за возбуждением"развивается в нейронах непосредственно после потенциала действия и характерно для клеток с длительной следовой гиперполяризацией. Таким образом, процессы торможения в локальных нейронных сетях уменьшают избыточную активность и участвуют в поддержании оптимальных режимов активности нейронов.
Механизмы координации рефлекторной деятельности: реципрокная иннервация, доминанта (А.А.Ухтомский), принципы обратной связи и общего конечного пути, принцип субординации.
Принцип иррадиации возбуждения. Иррадиация - распространение, расширение рефлекторного ответа. Это феномен “растекания” возбуждения по нейронам центральной нервной системы, развивающийся или после действия сверхсильного раздражителя, или на фоне выключения торможения. Распространение возбуждения возможно за счет многочисленных контактов между нейронами, возникающих при ветвлении аксонов и дендритов вставочных нейронов. Иррадиация позволяет увеличивать количество участвующих в рефлекторном ответе групп мышц. Ограничивают иррадиацию тормозные нейроны и синапсы.
На фоне действия стрихнина, блокирующего тормозные синапсы, наступают генерализованные судороги при тактильной стимуляции любого участка тела или при раздражении рецепторов любой сенсорной системы. В коре больших полушарий наблюдается явление иррадиации процесса торможения.
В основе координации рефлекторных актов лежат определенные механизмы, основанные на структурно-функциональной организации ЦНС и обозначаемые как “принципы” формирования рефлекторного ответа.
Принцип реципрокной иннервации. Реципрокная (сопряженная) координация открыта Н.Е.Введенским в 1896 году. Обусловлена реципрокным торможением, т.е. активация одного рефлекса одновременно сопровождается торможением второго, противоположного по своей физиологической сущности.
Принцип общего "конечного пути". Открыт английским физиологом Ч.Шеррингтоном (1906). Один и тот же рефлекс (например, сокращение мышцы) может быть вызван раздражением различных рецепторов, т.к. один и тот же конечный a-мотонейрон передних рогов спинного мозга входит в состав многих рефлекторных дуг. Рефлексы, дуги которых имеют общий конечный путь, подразделяются на агонистические и антагонистические. Первые усиливают, вторые тормозят друг друга, как бы конкурируя за конечный результат. В основе подкрепления лежит конвергенция и суммация, в основе конкуренции за конечный путь - сопряженное торможение.
Принцип обратной связи. Любой рефлекторный акт контролируется благодаря обратной связи с центром. Обратная связь состоит во вторичной афферентации, поступающей в ЦНС от рецепторов, которые возбуждаются при изменении функциональной активности рабочего органа. Например, потенциалы действия, обусловленные возбуждением рецепторов мышц, сухожилий и суставных сумок сгибающейся конечности, в процессе осуществления акта сгибания поступают во все структуры ЦНС, начиная от центров спинного мозга. Различают обратную связь положительную (усиливающую рефлекс, который является источником обратной афферентации) и отрицательную, когда рефлекс, ее вызывающий, тормозится. Обратная связь лежит в основе саморегуляции функций организма.
Принцип отдачи. Феномен отдачи состоит в быстрой смене одного рефлекса другим противоположного значения. Например, после сгибания конечности ее разгибание происходит быстрее, особенно если сгибание было сильным. Механизм этого явления состоит в том, что при сильном сокращении мышц возбуждаются рецепторы Гольджи сухожилий, которые через тормозные вставочные нейроны тормозят мотонейроны сгибательных мышц и образуют ветвь, которая возбуждает центр мышц - разгибателей. Благодаря этому механизму можно получить сумму рефлексов - цепные рефлексы (окончание одного рефлекторного ответа инициирует следующий) и ритмические (многократное повторение ритмичных движений).
Принцип доминанты. Конечный поведенческий эффект при координации рефлексов может быть изменен в зависимости от функционального состояния центров (наличия доминантных очагов возбуждения).
Особенности доминантного очага возбуждения:
1. Повышенная возбудимость нейронов.
2. Стойкость процесса возбуждения.
3. Способность к суммации возбуждения.
4. Иннертность. Очаг доминирует, подавляет соседние центры путем сопряженного торможения, возбуждаясь за их счет. Доминанту можно получить химическим воздействием на центры, например, стрихнином. В основе доминантного возбуждения лежит способность возбудительного процесса к иррадиации по нейронным цепям.
Торможение – активный, срочный процесс прекращения или задержки текущей функции под действием раздражителя.
Процесс торможения в центральной нервной системе был открыт в 1862 г. И. М. Сеченовым. В опытах на лягушках он делал поперечные разрезы головного мозга на различных уровнях и раздражал нервные центры, накладывая на разрез кристаллик поваренной соли. При этом обнаруживалось, что при раздражении промежуточного мозга наступает угнетение или полное торможение спинномозговых рефлексов: лапка лягушки, погруженная в слабый раствор серной кислоты, не отдергивалась.
Постсиналтическое торможение — основной вид торможения, развивающийся в постсинаптической мембране аксосоматических и аксодендритических синапсов под влиянием активации тормозных нейронов, в концевых разветвлениях аксонных отростков которых освобождается и поступает в синаптическую щель тормозной медиатор. Тормозной эффект таких нейронов обусловливается специфической природой медиатора — химического переносчика сигнала с одной клетки на другую. Наиболее распространенным тормозным медиатором является гамма-аминомасляная кислота (ГАМК). Химическое действие ГАМК вызывает в постсинаптической мембране эффект гиперполяризации в виде тормозных постсинаптических потенциалов (ТПСП), пространственно-временная суммация которых повышает уровень мембранного потенциала (гиперполяризация), приводит к урежению или полному прекращению генерации распространяющихся ПД.

А – возбуждающее окончание;Б – нейрон, вызывающий пресинаптическое торможение; В – нейрон, вызывающий постсинаптическое торможение.
Пресинаптическое торможение возникает перед синаптическим контактом – в пресинаптической области. Окончание аксона одной нервной клетки (терминаль) образует синапс на окончании аксона другой нервной клетки и блокирует передачу возбуждения в последней. В области такого пресинаптического контакта развивается чрезмерно сильная деполяризация мембраны аксона, которая приводит к угнетению проходящих здесь потенциалов действия.
Пресинаптическое торможение осуществляют нейроны (Б), аксоны которых оканчиваются на возбуждающих синаптических окончаниях другого нейрона (А), образуя аксо-аксональные синапсы (рис. 1Б). Пресинаптическое торможение выполняется посредством одного из двух механизмов.
1. Пресинаптический тормозным нейрон Б выделяет нейромедиатор, который увеличивает Cl–проводимость и вызывает гиперполяризацию мембраны возбуждающего нервного окончания А. Это приводит к снижению возбудимости и увеличению порога генерации ПД возбуждающего окончания.
2. Пресинаптический возбуждающий нейрон выделяет нейромедиатор, вызывающий длительную деполяризацию мембраны, что приводит к инактивациипотенциалозависимыхNa+-каналов нервного окончания и, соответственно, торможению его активности.
Координация - процессы взаимодействия между нейронами и нервным центром, которое обеспечивает их согласованную деятельность.
Свойство доминанта:1. повышенная возбудимость2. стойкость доминанты: могут сохранять часы, месяца, годы, всю жизнь 3.Инерция доминанты –последствие. 4. Способность к суммации
Сущность его заключается в том, что среди множества возбужденных очагов в ЦНС, существует очаг господствующего возбуждения. Этот доминантный очаг подчиняет себе все другие очаги возбуждения.
Принцип обратной связи. Всякий двигательный акт, вызванный афферентным раздражителем, сопровождается возбуждением рецепторов мышц, сухожилий, суставных сумок. Эти сигналы с проприорецепторов поступают в ЦНС. Обратная связь позволяет соотнести выраженность изменений параметров системы с ее работой. Связь выхода системы с ее входом с положительным коэффициентом усиления называется положительной обратной связью, а с отрицательным коэффициентом — отрицательной обратной связью. Положительная обратная связь в основном характерна для патологических ситуаций. Отрицательная обратная связь обеспечивает устойчивость системы. Механизмы обратной связи обеспечивают поддержание всех констант гомеостаза.
Принцип реципрокности. Он отражает характер отношение между центрами, ответственными за осуществление противоположных функций (вдоха и выдоха, сгибания и разгибания конечностей), и заключается в том, что нейроны одного центра, возбуждаясь, тормозят нейроны другого и наоборот.
Принцип временной связи. Высшая форма временной связи - условный рефлекс.
Вопрос 34.6. Роль спинного мозга в процессах регуляции деятельности опорно-двигательного аппарата и вегетативных функций организма. Рефлекторная деятельность спинного мозга. Клинически важные рефлексы.
Функции спинного мозга. Спинной мозг выполняет 2 основные функции: рефлекторную и проводниковую. Рефлекторная функция спинного мозга обеспечивает движение. Через спинной мозг проходят рефлекторные дуги, с которыми связано сокращение скелетных мышц тела (кроме мышц головы). Пример простейшего двигательного рефлекса — коленный рефлекс . Он проявляется в быстром подъеме ноги при резком ударе по сухожилию ниже коленной чашечки.
Спинной мозг вместе с головным мозгом регулирует работу внутренних органов: сердца, желудка, мочевого пузыря, половых органов. Белое вещество спинного мозга обеспечивает связь и согласованную работу всех отделов центральной нервной системы, осуществляя проводниковую функцию. Нервные импульсы, поступающие в спинной мозг от рецепторов, передаются по восходящим проводящим путям в головной мозг. Из головного мозга импульсы по нисходящим проводящим путям поступают к нижележащим отделам спинного мозга и оттуда — к органам.
Головной мозг регулирует работу спинного мозга. Известны случаи, когда в результате ранения или перелома позвоночника у человека прерывается связь между спинным и головным мозгом. Головной мозг у таких людей функционирует нормально. Но большинство спинномозговых рефлексов, центры которых расположены ниже места повреждения, исчезают. Такие люди могут поворачивать голову, совершать жевательные движения, изменять направление взгляда, иногда у них действуют руки. В то же время нижняя часть их тела лишена чувствительности и неподвижна.
Спинной мозг выполняет,.две главные функции — рефлекторную и проводниковую. Как рефлекторный центр спинной мозг способен осуществлять сложные двигательные и вегетативные рефлексы. Афферентными (чувствительными) путями спинной мозг связан с рецепторами, а эфферентными — со скелетной мускулатурой и со всеми внутренними органами. Длинные нисходящие и восходящие пути спинного мозга соединяют периферические части тела с головным мозгом.
По внешнему виду спинной мозг представляет собой продолговатый, несколько плоский цилиндрический тяж. Он расположен в позвоночном канале и на уровне нижнего края большого затылочного отверстия переходит в головной мозг.
Нижняя граница спинного мозга соответствуетуровнюI— II поясничных позвонков. Ниже этого уровня он продолжается в тонкую терминальную (концевую) нить.
У взрослого человека длина спинного мозга в среднем составляет около 43 см (у мужчин 45 см, у женщин 41 — 42 см), масса — около 34—38 г.
Функциональное разнообразие нейронов спинного мозга, наличие в нем афферентных нейронов, интернейронов, мотонейронов и нейронов автономной нервной системы, а также многочисленных прямых и обратных, сегментарных, межсегментарных связей и связей со структурами головного мозга — все это создает условия для рефлекторной деятельности спинного мозга с участием как собственных структур, так и головного мозга. Подобная организация позволяет реализовывать все двигательные рефлексы тела, диафрагмы, мочеполовой системы и прямой кишки, терморегуляции, сосудистые рефлексы и т. д.
Рефлекторные реакции спинного мозга зависят от места, силы раздражения, площади раздражаемой рефлексогенной зоны, скорости проведения по афферентным и эфферентным волокнам и, наконец, от влияния головного мозга. Сила и длительность рефлексов спинного мозга увеличивается при повторении раздражения
Сегментарная рефлекторная дуга состоит из рецептивного поля, из которого импульсация по чувствительному волокну нейрона спинального ганглия, а затем по аксону этого же нейрона через задний корешок входит в спинной мозг, далее аксон может идти прямо к мотонейрону переднего рога, аксон которого подходит к мышце. Так образуется моносинаптическая рефлекторная дуга, которая имеет один синапс между афферентным нейроном спинального ганглия и мотонейроном переднего рога.
Рассмотрим некоторые рефлексы спинного мозга
Миотатические рефлексы — рефлексы на растяжение мышцы. Быстрое растяжение мышцы, всего на несколько миллиметров механическим ударом по ее сухожилию приводит к сокращению всей мышцы и двигательной реакции. Например, легкий удар по сухожилию надколенной чашечки вызывает сокращение мышц бедра и разгибание голени. Дуга этого рефлекса следующая: мышечные рецепторы четырехглавой мышцы бедра à спинальный ганглий à задние корешки à задние рога III поясничного сегмента à мотонейроны передних рогов того же сегмента à экстрафузальные волокна четырехглавой мышцы бедра. Реализация этого рефлекса была бы невозможна, если бы одновременно с сокращением мышц-разгибателей не расслаблялись мышцы-сгибатели. Рефлекс на растяжение свойствен всем мышцам, но у мышц-разгибателей, они хорошо выражены и легко вызываются.
Висцеромоторные рефлексы возникают при стимуляции афферентных нервов внутренних органов и характеризуются появлением двигательных реакций мышц грудной клетки и брюшной стенки, мышц разгибателей спины.
Рефлексы автономной нервной системы имеют свои пути. Они начинаются от различных рецепторов, входят в спинной мозг через задние корешки, задние рога, далее в боковые рога, нейроны которых через передний корешок посылают аксоны не непосредственно к органам, а к ганглию симпатического или парасимпатического отдела автономной нервной системы.
Автономные (вегетативные) рефлексы обеспечивают реакцию внутренних органов, сосудистой системы на раздражение висцеральных, мышечных, кожных рецепторов. Эти рефлексы отличаются большим латентным периодом (ЛП) двумя фазами реакции: первая — ранняя — возникает с ЛП 7—9 мс и реализуется ограниченным числом сегментов, вторая — поздняя — возникает с большим ЛП — до 21 мс и вовлекает в реакцию практически все сегменты спинного мозга. Поздний компонент вегетативного рефлекса обусловлен вовлечением в него вегетативных центров головного мозга.
Читайте также:
