
Торможение внешней и внутренней нервной системы
Регуляция нервной деятельности представляет собой процессы возбуждения и торможения в ЦНС. Вначале она возникает как элементарная реакция на раздражение. В процессе эволюции произошло усложнение нейрогуморальных функций, приводящее к образованию основных отделов нервной и эндокринной систем. В данной статье мы изучим один из главных процессов – торможение в ЦНС, виды и механизмы его осуществления.
Нервная ткань, её строение и функции
Одна из разновидностей животных тканей, названная нервной, имеет особое строение, обеспечивающее как процесс возбуждения, так и приводящее в действие функции торможения в ЦНС. Нервные клетки состоят из тела и отростков: коротких (дендритов) и длинного (аксона), который обеспечивает передачу нервных импульсов от одного нейроцита к другому. Окончание аксона нервной клетки контактирует с дендритами следующего нейроцита в местах, называемых синапсами. Они обеспечивают передачу биоэлектрических импульсов по нервной ткани. Причем возбуждение всегда движется в одном направлении – с аксона на тело или дендриты другого нейроцита.
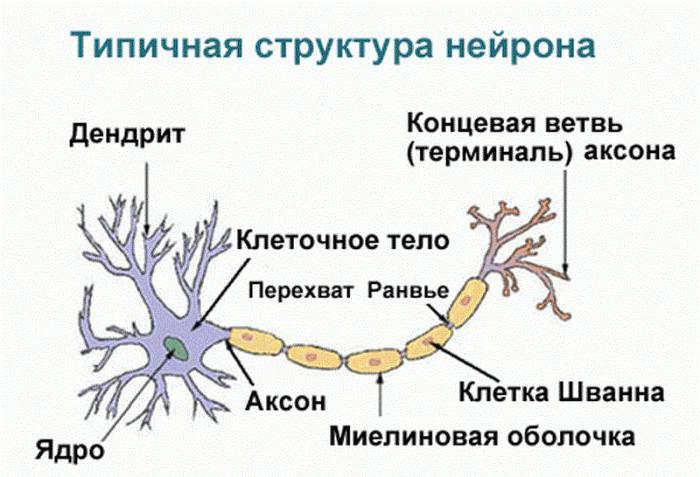
Еще одно свойство, кроме возбуждения, протекающее в нервной ткани, – торможение в ЦНС. Оно является ответной реакцией организма на действие раздражителя, ведущей к снижению или полному прекращению двигательной или секреторной активности, в которой участвуют центробежные нейроны. Торможение в нервной ткани может возникать и без предварительного возбуждения, а только лишь под воздействием тормозного медиатора, например ГАМК. Он является одним из главных трансмиттеров торможения. Здесь же можно назвать такое вещество, как глицин. Эта аминокислота участвует в усилении тормозных процессов и стимулирует в синапсах выработку молекул гаммааминомаслянной кислоты.
И. М. Сеченов и его работы в нейрофизиологии
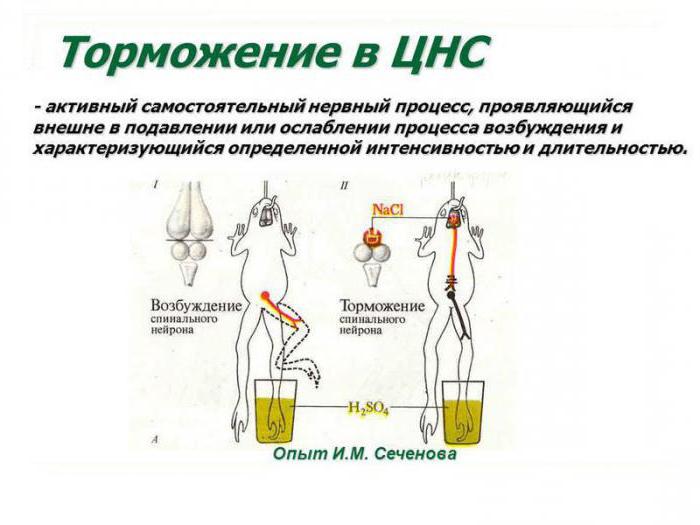
Выдающийся российский ученый, создатель теории рефлекторной деятельности головного мозга доказал наличие в центральных отделах нервной системы особых комплексов клеток, способных к инактивации биоэлектрических процессов. Открытие центров торможения в ЦНС стало возможным благодаря применению И. Сеченовым трех видов экспериментов. К ним относятся: перерезание участков коры в различных зонах головного мозга, стимуляция отдельных локусов серого вещества физическими или химическими факторами (электрическим током, раствором хлорида натрия), а также метод физиологического возбуждения мозговых центров. И. М. Сеченов был прекрасным экспериментатором, проводя сверхточные разрезы в зоне между зрительными буграми и непосредственно в самом таламусе лягушки. Он наблюдал уменьшение и полное прекращение двигательной активности конечностей животного.
Взаимодействуют ли нервные процессы?
Исследования выдающихся российских физиологов И. П. Павлова и И. М. Сеченова доказали, что работа центральной нервной системы характеризуется координацией рефлекторных реакций организма. Взаимодействие процессов возбуждения и торможения в ЦНС приводит к согласованной регуляции функций организма: двигательной активности, дыхания, пищеварения, выделения. Биоэлектрические процессы одновременно происходят в нервных центрах и могут последовательно меняться во времени. Это обеспечивает корреляцию и своевременное прохождение ответных рефлексов на сигналы внутренней и внешней среды. Многочисленные опыты, проведенные нейрофизиологами, подтвердили тот факт, что возбуждение и торможение в ЦНС – это ключевые нервные явления, в основе которых лежат некоторые закономерности. Остановимся на них подробнее.
Нервные центры коры головного мозга способны распространять оба вида процессов по всей нервной системе. Это свойство называется иррадиацией возбуждения или торможения. Противоположное явление – уменьшение или ограничение участка мозга, распространяющего биоимпульсы. Оно названо концентрацией. Оба вида взаимодействий ученые наблюдают в течение образования условных двигательных рефлексов. Во время начальной стадии формирования двигательных навыков, вследствие иррадиации возбуждения одновременно сокращаются сразу несколько групп мышц, не обязательно участвующих в выполнении формируемого двигательного акта. Только после многократных повторений формируемого комплекса физических движений (катания на коньках, лыжах, велосипеде), в результате концентрации процессов возбуждения в конкретных нервных очагах коры, все движения человека становятся высококоординированными.

Переключения в работе нервных центров могут происходить также вследствие индукции. Она проявляется при выполнении следующего условия: сначала происходит концентрация торможения или возбуждения, причем эти процессы должны быть достаточной силы. В науке известны два вида индукции: S-фаза (центральное торможение в ЦНС усиливает возбуждение) и отрицательная форма (возбуждение вызывает процесс торможения). Встречается также последовательная индукция. В этом случае нервный процесс меняется на противоположный в самом нервном центре. Исследования нейрофизиологов доказали тот факт, что поведение высших млекопитающих и человека определяется явлениями индукции, иррадиации и концентрации нервных процессов возбуждения и торможения.
Безусловное торможение
Рассмотрим более подробно виды торможения в ЦНС и остановимся на такой его форме, которая присуща как животным, так и человеку. Сам термин был предложен И. Павловым. Ученый считал этот процесс одним из врождённых свойств нервной системы и выделил два его вида: гаснущее и постоянное. Остановимся на них детальнее.
Допустим, в коре существует очаг возбуждения, генерирующий импульсы к рабочему органу (к мышцам, секреторным клеткам желез). Вследствие изменения условий внешней или внутренней среды возникает еще один возбужденный участок коры головного мозга. Он вырабатывает биоэлектрические сигналы большей интенсивности, что тормозит возбуждение в ранее активном нервном центре и его рефлекторной дуге. Гаснущее торможение в ЦНС приводит к тому, что интенсивность ориентировочного рефлекса постепенно уменьшается. Объяснение этому следующее: первичный раздражитель уже не вызывает процесса возбуждения в рецепторах афферентного нейрона.
Другой вид торможения, наблюдаемого как у человека, так и у животных, демонстрирует опыт, проведенный лауреатом нобелевской премии в 1904 году И. П. Павловым. Во время кормления собаки (с выведенной из щеки фистулой) экспериментаторы включали резкий звуковой сигнал – выделение слюны из фистулы прекращалось. Такой вид торможения ученый назвал запредельным.
Являясь врождённым свойством, торможение в ЦНС протекает по безусловно-рефлекторному механизму. Оно достаточно пассивно и не вызывает расхода большого количества энергии, приводя к прекращению условных рефлексов. Постоянное безусловное торможение сопровождает многие психосоматические заболевания: дискинезии, спастический и вялый параличи.
Что такое гаснущий тормоз
Продолжая изучать механизмы торможения в ЦНС, рассмотрим, что представляет собой один из его видов, названный гаснущим тормозом. Хорошо известно, что ориентировочный рефлекс представляет собой реакцию организма на воздействие нового постороннего сигнала. В этом случае в коре мозга образуется нервный центр, находящийся в состоянии возбуждения. Он и формирует рефлекторную дугу, отвечающую за реакцию организма и называемую ориентировочным рефлексом. Этот рефлекторный акт вызывает торможение условного рефлекса, происходящего в данный момент. После многоразового повторения постороннего раздражителя рефлекс, называемый ориентировочным, постепенно снижается и наконец исчезает. А значит, не вызывает больше торможения условного рефлекса. Такой сигнал и получил название гаснущего тормоза.
Таким образом, внешнее торможение условных рефлексов связано с влиянием на организм постороннего сигнала и является врождённым свойством центральной и периферической нервной системы. Внезапный или новый раздражитель, например, болевое ощущение, посторонний звук, изменение освещенности, не только вызывает ориентировочный рефлекс, но также способствует ослаблению или даже полному прекращению условно-рефлекторной дуги, активной в данный момент. Если посторонний сигнал (кроме болевого) действует повторно, торможение условного рефлекса проявляется меньше. Биологическая роль безусловной формы нервного процесса заключается в проведении ответной реакции организма на раздражитель, наиболее важный в данный момент.
Внутреннее торможение
Его другое название, используемое в физиологии высшей нервной деятельности, – условное торможение. Главная предпосылка возникновения такого процесса – отсутствие подкрепления сигналов, поступающих из внешнего мира, врождёнными рефлексами: пищеварительным, слюноотделительным. Возникшие в этих условиях процессы торможения в ЦНС требуют определенного временного интервала. Рассмотрим их виды более подробно.
Например, дифференцировочное торможение возникает как ответ на сигналы окружающей среды, совпадающие по амплитуде, интенсивности и силе к условному раздражителю. Эта форма взаимодействия нервной системы и окружающего мира позволяет организму более тонко различать раздражители и вычленять из их совокупности тот, который получает подкрепление врожденным рефлексом. Например, на звук звонка с силой 15 Гц, подкрепленный кормушкой с пищей, у собаки выработали условную слюноотделительную реакцию. Если к животному применить еще один звуковой сигнал, силой 25 Гц, не подкрепляя его пищей, в первой серии опытов у собаки из фистулы слюна будет выделяться на оба условных раздражителя. Через некоторое время у животного произойдет дифференциация этих сигналов, и на звук, силой 25 Гц слюна из фистулы перестанет выделяться, то есть разовьется дифференцировочное торможение.
Освободить мозг от информации, потерявшей жизненно значимую роль для организма, – эту функцию как раз и выполняет торможение в ЦНС. Физиология опытным путем доказала, что условные двигательные реакции, хорошо закрепленные выработанными навыками, могут сохраняться на протяжении всей жизни человека, например, катание на коньках, езда на велосипеде.
Подводя итог, можно сказать, что процессы торможения в ЦНС – это ослабление или прекращение определенных реакций организма. Они имеют очень большое значение, так как все рефлексы организма корригируются в соответствии с измененными условиями, а если условный сигнал потерял свое значение, то даже полностью могут исчезать. Различные виды торможения в ЦНС являются базовыми для таких способностей психики человека, как сохранение самообладания, различение раздражителей, ожидание.
Запаздывающий вид нервного процесса
Опытным путем можно создать ситуацию, при которой ответ организма на условный сигнал из внешней среды проявляется еще до воздействия безусловного раздражителя, например пищи. При увеличении промежутка времени между началом воздействия условного сигнала (свет, звук, например, удары метронома) и моментом подкрепления до трех минут выделение слюны на вышеназванные условные раздражители все более запаздывает и проявляется только в момент, когда перед животным появляется кормушка с пищей. Отставание ответа на условный сигнал характеризует процессы торможения в ЦНС, названные запаздывающим видом, при котором его время протекания соответствует интервалу запаздывания безусловного раздражителя, например пищи.
Значение торможения в ЦНС
Биологические основы поведения человека
Оба вида активности нервной ткани, как возбуждение, так и торможение в ЦНС, являются основой высшей нервной деятельности. Она обуславливает физиологические механизмы психической деятельности человека. Учение высшей нервной деятельности было сформулировано И. П. Павловым. Современная его трактовка звучит следующим образом:
- Возбуждение и торможение в ЦНС, происходящие во взаимодействии, обеспечивают сложные психические процессы: память, мышление, речь, сознание, а также формируют сложные поведенческие реакции человека.
Чтобы составить научно обоснованный режим учебы, труда, отдыха, ученые применяют знания закономерностей высшей нервной деятельности.
Биологическое значение такого активного нервного процесса, как торможение, можно определить следующим образом. Изменение условий внешней и внутренней среды (отсутствие подкрепления условного сигнала врождённым рефлексом) влечет за собой адекватные изменения приспособительных механизмов в организме человека. Поэтому приобретенный рефлекторный акт угнетается (гаснет) или вовсе исчезает, так как становится для организма нецелесообразным.
Что такое сон?
И. П. Павлов в своих работах экспериментально доказал тот факт, что процессы торможения в ЦНС и сон имеют единую природу. В период бодрствования организма на фоне общей активности коры головного мозга все же диагностируются отдельные её участки, охваченные внутренним торможением. Во время сна оно иррадиирует по всей поверхности больших полушарий, достигая подкорковых образований: зрительных бугров (таламуса), гипоталамуса, ретикулярной формации и лимбической системы. Как указывал выдающийся нейрофизиолог П. К. Анохин, все вышеперечисленные части центральной нервной системы, ответственные за поведенческую сферу, эмоции и инстинкты, во время сна снижают свою активность. Это влечет за собой снижение генерирования нервных импульсов, поступающих из-под корки. Таким образом, активизация коры снижается. Это обеспечивает возможность покоя и восстановления обмена веществ как в нейроцитах большого мозга, так и во всем организме в целом.

Опытами других ученых (Гесса, Экономо) были установлены особые комплексы нервных клеток, входящие в неспецифические ядра зрительных бугров. Процессы возбуждения, диагностируемые в них, вызывают снижение частоты биоритмов коры, которые можно расценивать как переход от активного состояния (бодрствования) ко сну. Исследования таких участков головного мозга, как Сильвиев водопровод и ІІІ желудочек, подтолкнули ученых к идее о наличии центра регуляции сна. Он анатомически связан с участком мозга, ответственным за бодрствование. Поражение этого локуса коры вследствие травмы или в результате наследственных нарушений у человека приводит к патологическим состояниям бессонницы. Также отметим тот факт, что регуляция такого жизненно важного для организма процесса торможения, как сон, осуществляется нервными центрами промежуточного мозга и подкорковых ядер: хвостатого, миндалевидного, ограды и чечевицеобразного.
Торможение — это нервный процесс, при котором возбуждение одних нейронов подавляет или предупреждает возбуждение других нейронов. Торможение никогда не распространяется по нервным волокнам.
Это местный процесс. Для торможения в ЦНС имеются специальные тормозные нейроны и тормозные синапсы. Некоторые медиаторы в ЦНС оказывают только тормозное действие, например, глицин, ГАМК, но есть медиаторы, которые оказывают как тормозное, так и возбуждающее действие, — ацетилхолин, глутамат.
Классификация торможения
Первичное
Постсинаптическое=гиперполяризационное
Пресинаптическое=деполяризационное
Торможение вслед за перевозбуждением (стрессом) → отключка.
1. Постсинаптическое, гиперполяризационное торможение связано с формированием на мембране ТПСП. Постсинаптическая мембрана при этом гиперполяризуется, возбудимость нейрона понижается, в результате он или уменьшает частоту, или прекращает генерировать ПД.
Виды постсинаптического торможения:
а) Поступательное опережающее торможение по Сеченову: при раздражении ретикулярной формации ствола мозга импульсы по ретикулоспинальному тракту поступают в спинной мозг и активируют тормозные интернейроны, которые тормозят альфа-мотонейроны. Торможение альфа-мотонейронов проявляется в увеличении латентного времени спинальных рефлексов.
б) Поступательное реципрокное торможение по Шеррингтону: при активации рецепторов растяжения какой-либо мышцы возбуждаются ее альфа-мотонейроны, при этом одновременно (реципрокно, сопряженно) тормозятся альфа-мотонейроны мышцы антагониста. Значение: автоматическое облегчение работы сустава.
в) Возвратное торможение по Реншоу: при увеличении частоты импульсации альфа-мотонейрона колатераль его аксона возбуждает тормозный интернейрон (клетка Реншоу). Тормозный нейрон выделяет медиатор глицин, который, вызывая процесс гиперполяризации, снижает величину деполяризации сомы альфа-мотонейрона, что приводит к понижению частоты ПД. Значение: обеспечивает сохранение средней частоты импульсации альфа-мотонейрона, нейрон импульсирует стабильно в течение длительного времени, обеспечивая поддержание полного тонуса.
г) Латеральное (окружающее) торможение: при активации одного нейрона одновременно активируются интернейроны, которые тормозят рядом лежащие нейроны. Это торможение хорошо изучено в сенсорных системах, предполагается, что оно обеспечивает контрастность восприятия сигнала.
2. Пресинаптическое деполяризационное торможение уменьшает эффект действия на клетку возбуждающих синапсов (торможение на входе). Морфологической основой его являются аксо-аксональные синапсы. Медиатор — ГАМК. При выделении ГАМК в пресинаптической терминали возбуждающего синапса развивается длительная деполяризация (до 100 мс), что приводит к уменьшению выделения возбуждающего медиатора и падению амплитуды ВПСП. Ионный механизм пресинаптического торможения до конца не выяснен. Предполагают, что происходит повышение проницаемости для ионов хлора, но двигается он согласно электрохимическому потенциалу из клетки.
Принципы взаимодействия нервных центров. Учение А.А. Ухтомского о доминанте.
Понятие о центральных двигательных программах.
Строение и связи нейронов нервного центра генетически детерминированы. За счет ветвления отростков и установления множества синапсов между клетками они образуют три основных типа нейронных сетей:
1. Иерархическая сеть распространяет афферентную информацию либо к увеличивающемуся число нейронов (принцип дивергенции), либо к меньшему числу нейронов (принцип конвергенции).
2. Локальная сеть – состоит из нейронов с короткими аксонами и обеспечивает взаимосвязь и сохранение информации в пределах одного уровня. Возбуждение в сети может циркулировать по замкнутому кольцу. Дублирование и взаимосвязанность элементов локальной сети обеспечивают надежность нервной регуляции.
3. Дивергентная сеть с одним входом – это нейронный ансамбль, который образует выходные связи с большим количеством разных НЦ. Сеть обеспечивает интеграцию разных рефлекторных актов и общую активность многочисленных нейронов разных отделов мозга.
Доминанта – это господствующий в данный момент очаг возбуждения в нервном центре, обуславливающий работу остальных нервных центров и определяющий направленность поведенческих реакций.
Доминанта — господствующий очаг возбуждения, предопределяющий характер текущих реакций центров в данный момент. Такой доминантный центр может возникнуть в различных этажах ЦНС при достаточно длительном действии гуморальных или рефлекторных раздражителей.
Физиологическую основу доминанты составляют отрицательная индукция и концентрация возбуждения. А сама доминанта является физиологической основой внимания, воли, восприятия и мышления. Принцип доминанты допускает, что если в коре мозга одновременно возникают два очага возбуждения, то один из них оказывается господствующим (доминирующим). Рефлексом, связанным с этим очагом в данный момент, направляется и трансформируется деятельность всего нервного аппарата.
Характеристика:
1. возбудимость обеспечивается таламусом
2. к недоминантным центрам оказывает тормозящее действие (осуществляют ядра таламуса)
3. доминантный очаг обладает стойкостью
4. наличие повышенной возбудимости
5. инерционность, обусловленная длительными следовыми процессами
6. способность к суммации и сопряженному торможению других центров, функционально не совместимых с деятельностью центров доминантного очага.
Способы смены доминанты:
ñ исчезает надобность
ñ появляется более сильная доминанта
Смену осуществляет лобная кора.
Дата добавления: 2018-08-06 ; просмотров: 538 ;
Павлов выделил два вида торможения условных рефлексов внешнее и внутреннее.
Высшая нервная деятельность основана на тончайшей координации функций больших полушарий и ближайших подкорковых отделов, осуществляемой благодаря взаимодействию двух форм нервного процесса: возбуждения и торможения. Нормальная высшая нервная деятельность происходит только в том случае, когда одновременно с возбуждением в больших полушариях и подкорковых центрах возникает и торможение. Торможение условных рефлексов имеет исключительное биологическое значение, так как оно обеспечивает соответствие условных рефлексов условиям существования и одновременно задерживает условные рефлексы, не имеющие или потерявшие свое значение для жизни.
Возбуждение и торможение — разные фазы в деятельности нейронов больших полушарий. Быстрое использование веществ во время возбуждения — главный толчок к появлению в нейроне торможения, которое не только ограничивает его дальнейшее функциональное разрушение, но и способствует восстановлению истраченных при возбуждении веществ. Биохимическая регенерация имеет огромное биологическое значение, так как таким образом обеспечивается сохранность и поддерживается способность к функционированию этих элементов организма, представляющих собой самую высокую ступень развития материи.
В больших полушариях различаются два вида торможения: внешнее и внутреннее.
Внешнее торможение — безусловное. Оно существует от рождения. Эта более простая форма торможения, выработавшаяся в процессе филогенетического развития, свойственна всей центральной нервной системе. В его осуществлении участвует ретикулярная формация.Внешнее торможение — результат взаимодействия соседних деятельных центров. Всякое новое внезапное раздражение, например посторонний звук, быстрое изменение освещения, появление струи воздуха и другие экстренные раздражители могут вызвать временное ослабление или даже полное уничтожение текущего условного рефлекса. Молодые, мало укрепившиеся условные рефлексы легче тормозятся, чем старые прочные условные рефлексы. Сильные посторонние раздражители тормозят условные рефлексы больше, чем слабые.
Внешнее торможение, как и в спинном мозге, — результат одновременной индукции. При действии постороннего раздражителя в соответствующей воспринимающей области возникает очаг возбуждения. А это приводит к тому, что одновременно в соседних областях, вследствие индукции, возникает торможение имеющихся в них более слабых очагов, возбуждения. Это торможение тем сильнее, чем сильнее очаг возбуждения, вокруг которого оно возникло. Внешнее торможение составляет физиологическую основу исчезновения концентрации внимания и его переключаемости.
Внешнее торможение может возникнуть и при отсутствии постороннего раздражителя. В этом случае оно появляется при действии очень сильного условного раздражителя, применяемого в данном опыте или при болевых раздражениях, и называется запредельным. Нейроны имеют предел работоспособности. При чрезмерной силе условного раздражителя, при сверхмаксимальном раздражении возникает опасность их разрушения и гибели. Торможение устраняет эту опасность и имеет охранительное значение. Предел работоспособности не является постоянной величиной и изменяется при утомлении, истощении, гипнотизации, заболеваниях, старости и т. д., а также зависит от функционального состояния нейронов. При нормальном или искусственном (путем введения химических веществ) повышении возбудимости нейронов все больше субмаксимальных или максимальных раздражителей превращается в сверхмаксимальные, и условно-рефлекторная деятельность понижается или прекращается. Это действие раздражителей сходно с явлением пессимума Н. Е. Введенского. Запредельное торможение относится к внешнему торможению, потому что оно возникает сразу без выработки и не тренируется, как внутреннее, условное торможение.
Внутреннее торможение — приобретенное, условное. Оно вырабатывается в течение индивидуальной жизни. Внутреннее торможение — непосредственный результат действия условных раздражителей. При внутреннем торможении положительные условные раздражители, вызывающие возбуждение, при определенных условиях превращаются в отрицательные условные раздражители, вызывающие торможение. В отличие от внешнего, внутреннее торможение непременно развивается постепенно и иногда вырабатывается очень медленно, с трудом.
Различают несколько разновидностей внутреннего торможения.
1. Угасательное торможение развивается при повторении условных раздражителей без подкрепления. Например, если звонок всегда сопровождается едой, он будет вызывать и движение животного к кормушке с пищей, и отделение слюны. Но если не кормить животное после этого звука, то звонок постепенно перестает вызывать двигательную и секреторную пищевые реакции. Скорость угасания зависит от частоты повторения условного раздражителя без подкрепления, от типа нервной системы, прочности ранее выработанного рефлекса и силы условного раздражителя. Она зависит также от интенсивности безусловного рефлекса, которым подкрепляется условный раздражитель, например от степени пищевой возбудимости, количества и качества пищи, применяемой для подкрепления. Таким образом, благодаря этой разновидности внутреннего торможения сигнал, который имел определенное жизненное значение, теперь не вызывает рефлекса. Не все условные рефлексы, образовавшиеся у животного при действии определенного раздражителя, угашаются одновременно. Например, после полного угашения секреторных и двигательных условных рефлексов долго не угасают (даже несколько лет) сердечные и дыхательные условные рефлексы — шизокинез. Особенно трудно поддаются угашению инструментальные оборонительные рефлексы. От угасательного торможения зависит степень приспособления животного к окружающей среде, так как оно регулирует поведение соответственно условиям жизни. Очевидно, способность избавления от ненужной привычки основана на этой разновидности внутреннего торможения.
Угасание происходит волнообразно. Оно тренируется по мере повторения опытов с угасанием на одном и том же животном. Торможение наступает раньше, и, наконец, полное угасание наступает после одного неподкрепления условного рефлекса. Угасание наблюдается не только на неподкрепленном рефлексе, но и на других подкрепляемых, что свидетельствует о распространении торможения. Через некоторое время угасший условный рефлекс восстанавливается самостоятельно, без подкрепления. Скорость его восстановления зависит от степени, глубины угасания, от типа нервной системы и от частоты повторения опытов с угасанием. Он восстанавливается также при действии экстренных посторонних раздражителей. Это явление обозначается как растормаживание.
2. Условное торможение. Для образования условного тормоза за несколько секунд до применения положительного условного раздражителя или одновременно с его действием прибавляют новый, ранее безразличный раздражитель, и эту комбинацию не подкрепляют. Постепенно условный раздражитель в соединении с новым раздражителем теряет свое положительное действие. Тот же условный раздражитель постоянно подкрепляется без присоединения нового раздражителя и продолжает вызывать условный рефлекс. Новый добавочный раздражитель становится условным тормозом, вызывающим торможение при присоединении его к любому положительному условному раздражителю.
Для образования условного тормоза необходимы следующие главные условия. Добавочный индифферентный раздражитель должен быть значительно сильнее основного положительного раздражителя. Интервал между прекращением действия добавочного раздражителя и основного положительного раздражителя должен быть при образовании условного тормоза на пищевой рефлекс меньше 10 с, или оба раздражителя должны применяться одновременно. При несоблюдении этих условий вырабатывается условный пищевой рефлекс второго порядка. При этих условиях можно образовать условный тормоз второго порядка. Скорость выработки условного торможения зависит от типа нервной системы животного, силы нового раздражителя и других условий. Условный тормоз тоже растормаживается при действии посторонних раздражителей. Условное торможение нужно отличать от внешнего торможения. Как и угасание, условное торможение образуется при неподкреплении. Условный тормоз по мере повторения действует все более короткое время. При подкреплении условного раздражителя условный тормоз угашается. Условным тормозом может стать и след от сильного раздражителя.
4. Дифференцировочное торможение. Условный рефлекс вызывается не только определенным условным раздражителем, но и близкими к нему раздражителями того же самого органа чувств.
Это обозначается как обобщение — генерализация. Генерализация условных рефлексов — физиологический процесс иррадиации возбуждения на соседние нейроны анализатора больших полушарий. Биологическое значение первоначальной генерализации раздражителей состоит в том, что в природе раздражители большей частью не оказываются строго определенными, а постоянно колеблются, меняются, переходя из одной родственной группы и в другую. Так, звук, издаваемый животным-врагом и являющийся условным раздражителем оборонительного рефлекса животного-жертвы, может колебаться по высоте, силе и тембру в зависимости от напряжения голосового аппарата, расстояния и т.д. Если один из этих близких раздражителей многократно не подкрепляется, то он постепенно перестает вызывать рефлекс и становится отрицательным условным раздражителем, а условный раздражитель, всегда подкрепляемый, продолжает вызывать рефлекс поэтому называется положительным. Например, условным раздражителем, вызывающим пищевую реакцию, сделан метроном, производящий 120 ударов в минуту. Сначала при действии метронома, производящего 140 или 100 ударов в минуту, также будет вызываться условный рефлекс. Но если метроном в 140 и 100 ударов никогда не подкреплять, то постепенно они перестанут вызывать условный рефлекс, а подкрепляемый метроном в 120 ударов будет вызывать рефлекс.
Следовательно, большие полушария головного мозга различают, дифференцируют близкие раздражители, действующие на один и тот же орган чувств (специализация условных рефлексов). Это различение обусловлено тем, что подкрепляемый условный раздражитель вызывает возбуждение, а неподкрепляемый дифференцировочный раздражитель — торможение. Дифференцировочное торможение в первый раз вырабатывается с трудом, но затем оно тренируется, так же как и другие разновидности внутреннего торможения. Поэтому процесс образования дифференцировочного торможения на другие условные раздражители все ускоряется, и новое дифференцировочное торможение вырабатывается быстрее. Дифференцировочное торможение вырабатывается с трудом, если сразу дифференцируется раздражитель, очень близкий по характеру к положительному условному раздражителю, и легче вырабатывается, если вначале дифференцируется далекий раздражитель, а затем уже отрицательный раздражитель все больше приближается к положительному. Чем интенсивнее положительный условный раздражитель, т. е. чем сильнее возбуждение, которому противостоит дифференцировочное торможение, тем оно более выражено. При повышении возбудимости больших полушарий, например при голодании, введении кофеина, ранее выработанное дифференцировочное торможение оказывается недостаточным. Дифференцировочное торможение также растормаживается при применении посторонних раздражителей. При внутреннем торможении нередко наблюдается расхождение секреторной и двигательной реакций: одна есть, а другой нет. Имеется торможение и при следовых рефлексах. Неподкрепляемые посторонние раздражители также могут вызвать торможение. Торможение легче, чем возбуждение, изменяется под влиянием посторонних раздражителей — тормозной процесс лабильнее.
Таким образом, в головном мозге существуют разнообразные формы торможения и его функция много сложнее, чем в нервных волокнах. Следует различать отсутствие рефлекса при недостаточной подпороговой силе условного или безусловного раздражителя или отсутствии условий его образования от активного задерживания, подавления условного или безусловного рефлексов.
Читайте также:
