
Трансформация стимула в нервную активность
Вторичночувствующие рецепторы отличаются от первичночувствующих механизмом трансформации стимула в нервную активность. Изменение электрического рецепторного потенциала высокоспециализированного рецептора под воздействием раздражителя приводит к выделению медиатора в область пресинаптической щели, расположенной между рецептором и окончанием нейрона. Вследствие изменения проницаемости постсинаптической мембраны нервных окончаний, подходящих к сенсорной клетке, появляется их деполяризация, которая приводит к развитию генераторного потенциала. Генераторный потенциал зависит целиком от внешнего стимула — его силы и длительности. Он стационарно удерживается в преобразующем участке нервного окончания и распространяется электротонически, с затуханием. Генераторный потенциал лишь при достижении порогового уровня запускает распространяющиеся импульсы сенсорного нейрона. Итак, преобразование энергии внешнего стимула и передача результатов этого преобразования в сенсорные ядра мозга обеспечивается двумя функционально различными процессами: градуальным генераторным потенциалом и потенциалом действия (импульсом), следующим закону "все или ничего".
По функциональным характеристикам рецепторы делят на монои полимодальные, спонтанноактивные и молчащие. Адаптация рецепторов — это снижение уровня их возбуждения под действием постоянно действующего раздражителя. Функциональное назначение рецепторов с различной степень адаптации: 1) слабо адаптирующиеся рецепторы служат для сигнализации об истинных, абсолютных и мгновенных величинах стимулов; 2) быстро и полностью адаптирующиеся рецепторы — для сигнализации об изменениях стимула на фоне шума — внешнего или биологического (пример внутреннего "шума" — спонтанная активность нейрона).
Когда рецепторы адаптируются к постоянно действующему стимулу, они теряют некоторое количество сведений о стимуле, например, о его продолжительности. Однако, чувствительность адаптированного рецептора к изменениям стимула возрастает. Любое усиление стимула действует на адаптированный рецептор как новый раздражитель.
3. Рецепторный потенциал.Рецепторный потенциалвозникает при действии внешнего стимула, который в результате появления ионных токов вызывает изменение потенциала покоя рецептора. Проницаемость мембраны рецептора к ионным токам, в основном, к токам Na + , в меньшей степени К + , Са 2+ , Cl меняется. Под действием стимула белковые молекулы белково-липидного слоя мембраны рецептора изменяют свою конфигурацию, и проводимость мембраны для мелких ионов повышается. Когда рецепторный потенциал достигает порогового значения, возникает нервный импульс — распространяющееся возбуждение. Такой рецепторный потенциал называют также генераторным потенциалом.
Рецепторный потенциал— изменение напряжения, возникающее в рецепторе при действии адекватного стимула вследствие изменения ионной проницаемости рецепторной мембраны, градуально зависящее от интенсивности стимула.
Рецепторный потенциал - это частный случай электротонических потенциалов. Когда рецепторная (сенсорная) клетка, например механочувствительная волосковая иливкусовая , подвергается воздействию соответствующего стимула, реализуется более или менее сложный набор событий, ведущих к изменениям электрической полярности участка их мембраны. Это явление именуется рецепторным потенциалом . В большинстве случаев рецепторные потенциалы - это деполяризация, в других, однако, в частности в палочках иколбочках сетчатки, - это гиперполяризация. Так или иначе, результат - одни и тот же - возникают токи между подвергающимся воздействию участком мембраны и другими участками мембраны рецепторной клетки. В общем случае, изменения электрической полярности (увеличение ее или уменьшение) влияет на выделение медиатора на подлежащий сенсорный нейрон.
Свойства рецепторных потенциалов:
1. Образуются в месте действия стимула.
2. Градуальность: стимулами разной интенсивности большинство рецепторов деполяризуется (или гиперполяризуется, как в случае палочек и колбочек) неодинаково. Хотя амплитуда потенциала определенным образом отражает силу стимуляции, последняя не служит источником энергии для такого изменения клетки. Единственная функция стимула - управление ионными токами через мембрану.
3. Локальность: распространяется по клетке электротонически, а не проводится активно по мембране.
4. Рецепторные потенциалы могут подвергаться пространственной и временной суммации.
Физиология восприятия
Общее представление
о восприятия стимула (раздражителя)
"Наше восприятие не воспроизводит окружающий мир, а создаёт его абстрактные модели".
Mountcastle V.B. The view from within: Pathways to the study of perception, John Hopkins Med. J. 136, # 3 (1975): 109-131 (p. 109).
Восприятие нервной системой чего-либо начинается с раздражения. Обязательно должен быть какой-то внешний по отношению к нервной системе раздражитель (стимул), который находится либо внутри, либо вне организм а, но обязательно вне нервной системы. Он должен воздействовать не на какую угодно точку организма или нервной системы, а на особые чувствительные клетки (сенсорные рецепторы), специально приспособленные для того, чтобы реагировать на него особенно легко, т.е. обладающие к нему повышенной чувствительностью. Только в таком случае воздействие физического фактора будет воспринято. Вот, к примеру, для восприятия радиации (радиоактивного излучения) у человеческого организма нет специальных сенсорных рецепторов, поэтому воздействие радиации не ощудается, несмотря на явный вред, который она может нанести организму.
В явлении восприятия раздражения следует понять одну простую, но важную вещь: сам стимул (раздражитель) находится снаружи и в нервную систему не проходит . По нервной системе бегает только возбуждение, порождённое этим стимулом, но не сам стимул!
Образно можно сказать об этом так:
В нервной системе нет ни света, ни цвета, ни звука, ни стука, ни вкуса, ни запаха, страдания и удовольствия в ней тоже нет — в ней есть одни только химические выбросы, электрические потенциалы и нервные импульсы! Но, к стати, знания и мысли в ней всё же есть!
Итак, восприятие раздражителя на уровне рецепторов заключается в переводе (трансформации) раздражения в нервное возбуждение . Это - одно из самых главных положений физиологии сенсорных систем.
Введём некоторые термины для лучшего понимания процесса восприятия: рецепция, трансдукция и перцепция. В большинстве случаев их можно применять как синонимы понятия "восприятие".
Рецепция - это первичное восприятие раздражения на уровне сенсорных рецепторов, приводящее к их возбуждению. Раздражение - это физическое воздействие подходящего (синоним: адекватного) раздражителя (синоним: стимула) на сенсорные рецепторы, которое вызывает их возбуждение. Возбуждение проявляется в виде локального (синоним: местного) изменения электрического потенциала клеточной мембраны.
Перцепция - это вторичное восприятие, т.е. построение нервной модели раздражения в виде сенсорного образа. Перцепция происходит на "верхних этажах" нервной системы, в то время как рецепция - на "нижних".
Результатом рецепции будет всего лишь поток нервных импульсов, поток возбуждения, а вот результатом перцепции уже будут субъективные ощущения, хотя и то, и другое, несомненно, можно назвать восприятием в целом.
Трансдукция (трансформация) - это преобразование раздражения в возбуждение сенсорными рецепторами.
Трансдукция = раздражение --> рецептор -->возбуждение.
Здесь важно помнить, что возникшее в сенсорных рецепторах возбуждение вовсе не обязательно должно иметь вид нервных импульсов. Как правило, оно имеет вид " рецепторного потенциала ", т.е. деполяризации мембраны. Есть и исключения: так, фоторецепторы реагируют на световое раздражение не деполяризацией, а наоборот, гиперполяризацией.
Вторичночувствующий сенсорный рецептор - это специализированная клетка, способная в ответ на раздражение создавать локальный рецепторный потенциал и выделять медиатор, воздействующий на окончание связанного с ней афферентного нейрона. А уже афферентный нейрон под воздействием медиатора порождает нервный импульс, т.е. распространяющееся возбуждение.
Зрительная система, в отличие от остальных сенсорных систем, работает особым образом. Фоторецепторы в ответ на раздражение развивают не деполяризацию, а, наоборот, гиперполяризацию и уменьшают выделение своего медиатора. Кроме того, связанные с фоторецепторами афферентные биполярные нейроны, подобно им, не умеют порождать нервные импульсы, а формируют локальные рецепторные потенциалы и выделяют медиатор.
В целом работа вторичночувствующих сенсорных рецепторов очень похожа на работу обычных синапсов, только запускается она раздражителем.
Общая схема трансдукции раздражения в нервное возбуждение в рецепторах вторичночувствующих
(это очень важно понять, так как рецепторы - это входы в нервную систему, только через них нервная система общается с остальным миром)
Первый этап : Раздражитель запускает в рецепторную клетку ионы Na + для создания деполяризации, т.е. смещения электрического потенциала мембраны в сторону нуля
1. Раздражитель прямо или опосредованно воздействует на сенсорный рецептор.
2. В ответ на это воздействие в мембране рецепторной клетки обязательно должны открыться ионные каналы для натрия ("стимул-управляемые" ионные каналы).
3. Через открытые натриевые каналы в клетку заходят ионы натрия Na + , приносят положительные заряды и вызывают деполяризацию мембраны. Это локальный рецепторный потенциал. Он не способен разбегаться по мембране или по отросткам клеток. Но он способен открыть те ионные каналы, на которые может действовать деполяризация.
Второй этап : Деполяризация запускает в рецепторную клетку ионы Ca 2+ для активирования пузырьков с медиатором
4. Деполяризация открывает ионные каналы для второго типа ионов - для ионов кальция Ca 2+ (не калия!).
5. Через открытые каналы кальций входит в клетку. Важно не то, что он приносит положительные заряды, увеличивая деполяризацию, а то, что он сам является мощным биологическим активатором и запускает процесс перемещения пузырьков с медиатором к мембране и выделение медиатора в синаптическую щель, за которой располагается воспринимающее окончание афферентного нейрона.
Третий этап : Медиатор запускает в афферентный нейрон ионы Na + для создания деполяризации (локального потенциала)
6. Медиатор, выделившийся из сенсорной рецепторной клетки, продвигается через синаптическую щель и связывается с молекулярными рецепторами нервного окончания афферентного нейрона.
7. Под действием медиатора на окончании афферентного нейрона открываются "хемоуправляемые" ионные каналы для натрия Na + .
8. Ионы натрия входят в окончание и вызывают деполяризацию мембраны, которая называется здесь "локальный потенциал". Таким образом возникает локальный потенциал на воспринимающем окончании афферентного нейрона.
Четвёртый этап : Деполяризация (локальный потенциал) на мембране афферентного нейрона порождает (генерирует) на нём нервный импульс
9. При достижении критического уровня деполяризации (порогового потенциала) на афферентном окончании открываются "потенциал-управляемые" ионные каналы для натрия Na + . Начинается самоусиливающийся поток ионов натрия в клетку. И волна изменений бежит по мембране аксона афферентного нейрона. А это уже - потенциал действия и нервный импульс !
10. Таким образом, нервное окончание афферентного нейрона, связанное с рецепторной клеткой, формирует генераторный потенциал на аксонном холмике афферентного нейрона, который при достижении критического уровня деполяризации переходит в потенциал действия и нервный импульс. Так рождается распространяющееся нервное возбуждение, которое можно назвать "сенсорным возбуждением".
Термин "трансдукция" означает "перенос". Он употребляется в этом значении не только в физиологии возбуждения, но и в других разделах биологии, где уже не связан с преобразованием раздражения в возбуждение.
В чём же заключается сущность восприятия в целом?
Сущность восприятия:
1. В ответ на раздражение нервная система порождает нервное возбуждение (нервные импульсы) — это сенсорное возбуждение.
Это и есть главные положения физиологии восприятия стимула (раздражителя).
Этапы восприятия раздражения (стимула)
Раздражение → сенсорные рецепторы : рецепция / трансдукция - трансформация раздражения в возбуждение / кодирование → поток сенсорного возбуждения → низшие нервные центры : детекция / перекодирование /трансформация / разделение/слияние/перенаправление потоков возбуждения → потоки сенсорного, эффекторного и модулирующего возбуждения → высшие нервные центры : создание сенсорного образа/модели / детекция/анализ/синтез.
После этого происходит направление возбуждения из высшего сенсорного нервного центра на управляющие и эффекторные системы.
Процесс восприятия также изображается в виде картинки на нашей схеме: Пути сенсорного возбуждения.
Адекватный стимул, классификация рецепторов.Рецептор-это специализированная нервная клетка. Она сигнализирует в ЦНС о состоянии и (или) изменениях состояния среды, в которой находится. Те факторы окружающей среды и те их изменения, которые оказывают действие на рецепторы, называются стимулами. Стимулы-это величины, которые можно измерять объективными методами, например деформация кожи, температура и электромагнитное излучение (свет).
Физиологические исследования обнаружили, что каждый рецептор с особой готовностью реагирует на стимулы какого-то одного типа. Это свойство обычно называют специфичностью рецепторов; тот стимул, который является эффективным в каждом случае, иногда называют адекватным стимулом для данного рецептора. Для некоторых рецепторов адекватный стимул можно установить из повседневного опыта. Так, например, мы можем легко определить, что адекватным стимулом для рецепторов наших глаз является свет, тогда как тепловые или механические стимулы с помощью этих рецепторов мы в обычных условиях не обнаруживаем.
Следовательно, каждый рецептор способен передавать в ЦНС информацию только об одном определенном аспекте или размерности окружающей среды. Это значит, что рецептор играет роль фильтра. В связи с этим представляется полезным классифицировать рецепторы на основе их адекватных стимулов. Рецепторы млекопитающих распадаются на следующие" четыре группы: механо-, термо-, хемо-и фоторецепторы.
Внутри каждой из этих четырех групп мы можем усмотреть значительную степень специализации. Например, есть различающиеся типы фоторецепторов, по-разному отвечающие на излучения с различными длинами
волн; эти рецепторы называют чувствительными к длине во
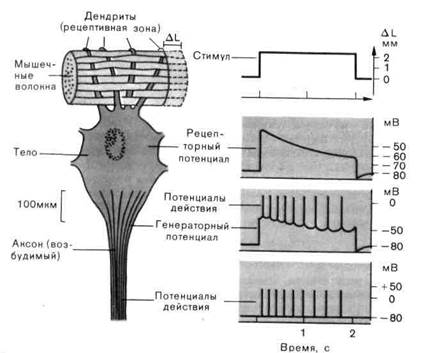
гас. г-1. схема электрофизиологических измерений, которые можно провести на рецепторе растяжения ракообразных. Во время стимуляции (изменение длины соответствующей мышцы на AL) микроэлектрод, введенный в тело клетки, регистрирует рецепторный потенциал, который возникает в рецептивной зоне (на дендритах). Являясь генераторным потенциалом, он возбуждает распространяющиеся потенциалы действия в возбудимом аксоне.
лны или цветочувствительными. Терморецепторы можно подразделить на рецепторы тепла и холода в соответствии с тем, что их возбуждает -увеличение или уменьшение температуры. У механорецепторов также можно наблюдать специализацию к различным параметрам стимулов: в коже, например, одни рецепторы чувствительны к вибрации, другие-к давлению.
Рецепторный потенциал. Вследующих параграфах мы рассмотрим процессы в рецепторе (подвергаемом действию адекватного стимула), результатом которых является потенциал действия в афферентном волокне. В качестве примера взят рецептор растяжения ракообразных-ме-ханорецептор, расположенный между определенными мышечными волокнами животного; он чувствителен к растяжению (рис. 2-1).
Когда микроэлектрод проникает в рецепторную клетку, то потенциал, который регистрируется исходно, как и во всех других нервных клетках, - это потенциал покоя (-80мВ на рис. 2-1). Под действием адекватной стимуляции - когда окружающая мышца растягивается мембранный потенциал меняется в направлении деполяризации. Эта вызываемая стимулом деполяризация-отклонение от потенциала покоя-называется рецепторным потенциалом. Рецепторный потенциал длится столько же, сколько и стимул (изменение ∆L на рис. 2-1). Однако даже тогда, когда стимул сохраняется постоянным, рецепторный потенциал снижается от своего исходного максимального значения до более низкого уровня. Такое снижение влияния стимула, поддерживаемого в течение некоторого времени на постоянном уровне, можно наблюдать практически у всех рецепторов; оно называется адаптацией.
Рецепторный потенциал в этом случае является результатом повышения проводимости мембраны, которое неспецифично и касается всех мелких ионов (Na + , K + , Са 2+ , Сl‾). В нормальных условиях единственный из этих ионов, который может обеспечить деполяризацию, это ион Na + , потому что только этот ион имеет потенциал равновесия, сдвинутый в деполяризационном направлении по отношению к потенциалу покоя. Следовательно, ионы Na + должны быть главным источником возникновения рецепторного потенциала. Повышение проводимости мембраны, описанное для рецепторов, сходно с тем, что имеет место в субсинаптической мембране возбуждающих синапсов - например в концевой пластинке мотонейрона.
Фоторецепторы позвоночных представляют исключение в том смысле, что при стимуляции светом в них возникает гиперполяризационный потенциал. Однако при синаптической передаче на нейроны высших уровней в сетчатке индуцируется деполяризация-возбуждающий пост-синаптический потенциал (ВПСП), который ведет к генерации распространяющихся потенциалов действия.
Процесс трансдукции; первичные и вторичные сенсорные клетки.Промежуточные события между поступлением стимула и появлением рецепторного потенциала пока еще мало изучены; они объединяются под общим названием процессы трансдукции. Этот термин подразумевает главным образом молекулярно-биологические процессы, имеющие место на рецептивной мембране и (или) во внутриклеточных структурах (называемых органеллами). В воспринимающей части рецептора имеется много таких органелл, как мембранные диски фоторецепторов, ко
 |
торые содержат чувствительные к свету молекулы зрительного пигмента (см. рис. 4-27) или волосковидные выросты (реснички) рецепторов внутреннего уха (см. рис. 5-3). Это вспомогательные структуры, которые делают рецептор особенно чувствительным к специфическому адекватному стимулу.
Многие рецепторы состоят из двух связанных элементов: нервного окончания и некоторой клетки ненервного происхождения типа дисков Меркеля в коже (см. рис. 3-4) или чувствительных клеток во вкусовых почках (см. рис. 7-3). Полагают, что процесс преобразования сигнала имеет место в не-нервной клетке. Такие клетки называют вторичными сенсорными клетками. На электронной микрофотографии область контакта между клетками выглядит как синапс. По этой причине предполагается, что имеется синаптическая передача от вторичной сенсорной клетки к нервному окончанию.
Напротив, первичная сенсорная клетка -это рецептор, в котором преобразование стимула происходит в нервной клетке или в нервном окончании. Примером этого являются обонятельные рецепторы (см. рис. 8-2) и свободные нервные окончания в коже.
Рецепторный потенциал как причина генерации распространяющихся потенциалов действия.Рецепторный потенциал электротонически распространяется в смежные области клетки. В результате этого аксон деполяризуется (рис. 2-1). Когда деполяризация достигает порога мембраны аксона, генерируется потенциал действия. Этот потенциал действия проводится по аксону в ЦНС. Таким образом, рецепторный потенциал действует на аксон как электрический стимул; по этой причине его также называют генераторным потенциалом.
Если генераторный потенциал не спадает после генерации первого потенциала действия, он может вызывать следующие потенциалы действия, пока стимул и соответственно генераторный потенциал не исчезнут. Это значит, что в период существования длительного генераторного потенциала афферентное волокно разряжается многократно. Процессы, происходящие между поступлением стимула и генерацией распространяющихся потенциалов действия, могут быть названы процессами трансформации стимула в нервную активность. Таким образом, эта трансформация включает следующие события: трансдукцию, увеличение проводимости рецептивной мембраны, рецепторный потенциал, синаптическую передачу (в случае вторичных сенсорных клеток), генераторный потенциал, потенциалы действия.
Адаптация. При рассмотрении рецепторного потенциала мы отметили, что даже если стимул поддерживается некоторое время постоянным, рецепторный потенциал может уменьшаться (рис. 2-1); этот процесс называют адаптацией. Мы также наблюдаем этот феномен на картине разряда афферентных нервных волокон: как можно видеть из рис. 2-1, при неизменном стимуле временные интервалы ∆t между двумя последовательными импульсами удлиняются, т.е. мгновенная частота разряда уменьшается (мгновенная частота = 1/∆t). Адаптация может иметь место на всех стадиях трансформации стимула: в процессе трансдукции, в механизме проведения рецепторного потенциала, при синаптической передаче от вторичных сенсорных клеток и при генерации потенциала действия. Возможно, что несколько таких механизмов работает одновременно.

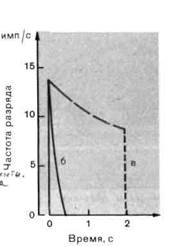
Рис. 2-2. Разряды рецепторов с различными скоростями адаптации. А. Потенциалы действия очень быстро (а), быстро (б) и медленно (в) адаптирующихся рецепторов, генерируемые при длительном действии постоянного стимула. Б. Графики изменения мгновенной частоты разряда со временем для рецепторов (б) и (в). Обратите внимание на то, что разряд рецептора (в) включает как фазиче-ский, так и тонический компоненты.
Чтобы получить адаптационную характеристику рецептора, исследуют его реакцию на надпороговый ступенчатый стимул. В таких экспериментах различные рецепторы демонстрируют различные изме
нения частоты разряда во времени. На рис. 2-2, А показан разряд медленно адаптирующегося рецептора (в), умеренно адаптирующегося рецептора (б) и быстро адаптирующегося рецептора (а). На рис. 2-2, Б представлено изменение со временем мгновенной частоты для рецепторов с медленной и умеренной адаптацией.
Предложены различные функциональные объяснения адаптации. Это явление обычно рассматривают всего лишь как простое утомление рецепторов. Но во многих случаях различное развитие адаптации во времени можно интерпретировать как приспособление к передаче специфических параметров стимула. Например, при изучении трех типов ме-ханорецепторов кожи было обнаружено, что благодаря характерным временным свойствам их разряда они могут рассматриваться как пере-
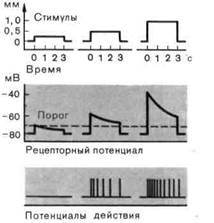 |
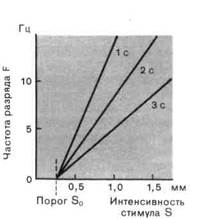 Рис. 2-3. Зависимость рецепторного потенциала и частоты разряда от интенсивности стимула. А. Рецепторные потенциалы и потенциалы действия при трех различных интенсивностях стимула. Потенциалы действия возбуждаются тогда, когда рецепторный потенциал превосходит порог (красная прерывистая линия). Б. Графики зависимости мгновенной частоты разряда F в разные моменты после включения стимула (через 1, 2 и 3 с) от интенсивности стимула S. S0 -пороговая интенсивность. |
 |
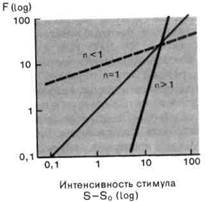 |
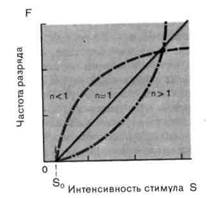
Рис. 2-4. Различные степенные функции. А. Частота разряда F как функция интенсивности стимула S. По обеим осям масштаб линейный, единицы-произвольные. Приведены три характерные степенные функции с показателями п 1 и п = 1 (красная линия). Б. Те же степенные функции изображены в логарифмическом масштабе. По абсциссе отложена разность между интенсивностью стимула S и пороговой интенсивностью S0.
датчики сведений об амплитуде, скорости и ускорении деформации кожи (см. рис. 3-5-3-8 и табл. 3-1).
Перевод интенсивности стимула в частоту импульсов в разряде.Когда сила стимула возрастает, возрастает и амплитуда рецепторного потенциала. Постепенные плавные изменения рецепторного потенциала вызывают соответствующие изменения частоты потенциалов действия. Эта ситуация представлена на рис. 2-3, А. Соотношение между интенсивностью стимула S и частотой импульсов F, часто называемое силовой функцией, изображено на рис. 2-3, £ для разных моментов времени от начала стимуляции (через 1, 2 и 3 с). В случае рецептора растяжения соотношение между F и S линейно для каждого момента времени на протяжении действия стимула.
Поскольку для того, чтобы вызвать потенциалы действия, рецепторный потенциал должен превзойти некоторое минимальное значение (порог аксона), разряд возникает только при превышении стимулом соответствующей интенсивности (S0 на рис. 2-3, Б). S0 называют пороговым стимулом для рецептора. Силовые функции рис. 2-3, Б описываются выражениями вида
т. е. линейной зависимостью для S > S0. Коэффициент к, наклон прямой, становится все меньше для последовательных интервалов времени, от-
считываемых от момента приложения стимула; это тоже является одним из выражений адаптации. '
Экспериментально было показано, что для большинства рецепторов силовая функция нелинейна. Измеренные соотношения часто можно описать различными математическими функциями, такими, как логарифмическая
Мы сталкивались с обоими этими выражениями в разделе по психофизике; они называются соответственно функциями Вебера-Фехнера иСтивенса. Кодирование интенсивности в рецепторах большинства типов лучше всего описывается степенной функцией (рис. 2-4), в которой разность- между интенсивностью стимула S и пороговой интенсивностью S0 возводится в n-ю степень. Показатель п является константой, характерной для каждого типа рецепторов. Для п 1 степенные функции (на графиках с линейным масштабом) представляются соответственно выпуклыми и вогнутыми кривыми (рис. 2-4, А), а для п = 1 силовая функция линейна, как и для рецептора растяжения. Оказалось, что для большинства рецепторов и + (значением рН);
д) медленно нарастает при действии постоянного стимула
и длится в течение всего времени стимуляции.
Верны несколько ответов.
В 2.3. В афферентных аксонах многих рецепторов частота импульсов в разряде
а) увеличивается с увеличением интенсивности стимула;
б) увеличивается со временем при действии стимула постоянной интенсивности;
в) уменьшается со временем при действии стимула постоянной
г) обычно равна нулю при действии стимулов подпороговой
д) не зависит от величины рецепторного потенциала. Верны несколько ответов.
В 2.4. В случае многих рецепторов степенная функция описывает
а) временной ход адаптации;
б) временной ход рецепторного потенциала при увеличении силы
стимула;
в) зависимость частоты импульсации F от величины превышения
стимулом порога S0;
г) связь между амплитудой потенциала действия и величиной ре
цепторного потенциала.
Дата добавления: 2015-10-05 ; просмотров: 1840 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Преобразование энергии раздражителя в рецепторах. Рецепторный потенциал. Абсолютный порог. Длительность ощущения. Адаптация рецепторов.
В результате действия адекватного раздражителя у большинства рецепторов увеличивается проницаемость клеточной мембраны для катионов, что приводит к ее деполяризации. Исключением из общего правила являются фоторецепторы, где после поглощения энергии квантов света в связи особенностями управления ионными каналами происходит гиперполяризация мембраны. Изменение величины мембранного потенциала рецепторов в ответ на действие стимула представляет собой рецепторный потенциал — входной сигнал первичных сенсорных нейронов. Если величина рецепторного потенциала достигнет критического уровня деполяризации или превысит его, генерируются потенциалы действия, с помощью которых сенсорные нейроны передают в центральную нервную систему информацию о действующих стимулах.
Генерация потенциалов действия происходит в ближайшем к рецепторам перехвате Ранвье миелинизированных волокон или ближайшей к рецепторам части мембраны безмиелинового волокна. Минимальная сила адекватного стимула, достаточная для генерации потенциалов действия в первичном сенсорном нейроне, определяется как его абсолютный порог. Минимальный прирост силы стимула, сопровождающийся значимым изменением реакции сенсорного нейрона, представляет собой дифференциальный порог его чувствительности.
Информация о силе действующего на рецепторы стимула кодируется двумя способами: частотой потенциалов действия, возникающих в сенсорном нейроне (частотное кодирование), и числом сенсорных нейронов, возбудившихся в ответ на действие стимула. При увеличении силы действующего на рецепторы раздражителя повышается амплитуда рецепторного потенциала, что, как правило, сопровождается увеличением частоты потенциалов действия в сенсорном нейроне первого порядка. Чем шире имеющийся частотный диапазон потенциалов действия у сенсорных нейронов, тем большее число промежуточных значений силы раздражителя способна различать сенсорная система. Первичные сенсорные нейроны одинаковой модальности различаются порогом возбуждения, поэтому при действии слабых стимулов возбуждаются только наиболее чувствительные нейроны, но с увеличением силы раздражителя на него реагируют и менее чувствительные нейроны, имеющие более высокий порог раздражения. Чем больше первичных сенсорных нейронов возбудится одновременно, тем сильнее будет их совместное действие на общий нейрон второго порядка, что в итоге отразится на субъективной оценке интенсивности действующего раздражителя.
Длительность ощущения зависит от реального времени между началом и прекращением воздействия на рецепторы, а также от их способности уменьшать или даже прекращать генерацию нервных импульсов при продолжительном действии адекватного стимула. При длительном действии стимула порог чувствительности рецепторов к нему может повышаться, что определяется как адаптация рецепторов. Механизмы адаптации не одинаковы в рецепторах разных модальностей, среди них различают быстро адаптирующиеся (например, тактильные рецепторы кожи) и медленно адаптирующиеся рецепторы (например, проприоцепторы мышц и сухожилий). Быстро адаптирующиеся рецепторы сильнее возбуждаются в ответ на быстрое нарастание интенсивности стимула (фазический ответ), а их быстрая адаптация способствует освобождению восприятия от биологически незначительной информации (например, контакт между кожей и одеждой). Возбуждение медленно адаптирующихся рецепторов мало зависит от скорости изменения стимула и сохраняется при его длительном действии (тонический ответ), поэтому, например, медленная адаптация проприоцепторов позволяет человеку получать нужную ему для сохранения позы информацию в течение всего необходимого времени.
Существуют сенсорные нейроны, генерирующие потенциалы действия спонтанно, т. е. при отсутствии раздражения (например, сенсорные нейроны вестибулярной системы), такая активность называется фоновой. Частота нервных импульсов в этих нейронах может увеличиваться или уменьшаться в зависимости от интенсивности действующего на вторичные рецепторы стимула, кроме того, она может определяться направлением, в котором отклоняются чувствительные волоски механорецепторов. Например, отклонение волосков вторичных механорецепторов в одну сторону сопровождается повышением фоновой активности сенсорного нейрона, которому они принадлежат, а в противоположную сторону — понижением его фоновой активности. Указанный способ рецепции позволяет получать информацию и об интенсивности стимула, и о направлении, в котором он действует.
Читайте также:
