
В образовании клеточных мембран оболочек нервных клеток участвуют
Клеточная мембрана также называется плазматической (или цитоплазматической) мембраной и плазмалеммой. Данная структура не только отделяет внутреннее содержимое клетки от внешней среды, но также входит с состав большинства клеточных органелл и ядра, в свою очередь отделяя их от гиалоплазмы (цитозоля) — вязко-жидкой части цитоплазмы. Договоримся называть цитоплазматической мембраной ту, которая отделяет содержимое клетки от внешней среды. Остальными терминами обозначать все мембраны.
Строение клеточной мембраны
В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.
В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков. Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.
Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Функции клеточной мембраны
Большинство погруженных в клеточную мембрану белков выполняют ферментативную функцию (являются ферментами). Часто (особенно в мембранах органоидов клетки) ферменты располагаются в определенной последовательности так, что продукты реакции, катализируемые одним ферментом, переходят ко второму, затем третьему и т. д. Образуется конвейер, который стабилизируют поверхностные белки, т. к. не дают ферментам плавать вдоль липидного бислоя.
Клеточная мембрана выполняет отграничивающую (барьерную) от окружающей среды и в то же время транспортную функции. Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).
При этом транспорт веществ происходит различными способами. Транспорт по градиенту концентрации предполагает передвижение веществ из области с их большей концентрацией в область с меньшей (диффузия). Так, например, диффундируют газы (CO2, O2).
Бывает также транспорт против градиента концентрации, но с затратой энергии.
Транспорт бывает пассивным и облегченным (когда ему помогает какой-нибудь переносчик). Пассивная диффузия через клеточную мембрану возможна для жирорастворимых веществ.
Есть особые белки, делающие мембраны проницаемыми для сахаров и других водорастворимых веществ. Такие переносчики соединяются с транспортируемыми молекулами и протаскивают их через мембрану. Так переносится глюкоза внутрь эритроцитов.
Пронизывающие белки, объединяясь, могут образовывать пору для перемещения некоторых веществ через мембрану. Такие переносчики не перемещаются, а образуют в мембране канал и работают аналогично ферментам, связывая определенное вещество. Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример — натрий-калиевый насос.
Транспортная функция клеточной мембраны эукариот также реализуется за счет эндоцитоза (и экзоцитоза). Благодаря этим механизмам в клетку (и из нее) попадают крупные молекулы биополимеров, даже целые клетки. Эндо- и экзоцитоз характерны не для всех клеток эукариот (у прокариот его вообще нет). Так эндоцитоз наблюдается у простейших и низших беспозвоночны; у млекопитающих лейкоциты и макрофаги поглощают вредные вещества и бактерии, т. е. эндоцитоз выполняет защитную функцию для организма.
Эндоцитоз делится на фагоцитоз (цитоплазма обволакивает крупные частицы) и пиноцитоз (захват капелек жидкости с растворенными в ней веществами). Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Экзоцитоз — это выведение цитоплазматической мембраной веществ из клетки (гормонов, полисахаридов, белков, жиров и др.). Данные вещества заключаются в мембранные пузырьки, которые подходят к клеточной мембране. Обе мембраны сливаются и содержимое оказывается за пределами клетки.
Цитоплазматическая мембрана выполняет рецепторную функцию. Для этого на ее внешней стороне располагаются структуры, способные распознавать химический или физический раздражитель. Часть пронизывающих плазмалемму белков с наружней стороны соединены с полисахаридными цепочками (образуя гликопротеиды). Это своеобразные молекулярные рецепторы, улавливающие гормоны. Когда конкретный гормон связывается со своим рецептором, то изменяет его структуру. Это в свою очередь запускает механизм клеточного ответа. При этом могут открываться каналы, и в клетку могут начать поступать определенные вещества или выводиться из нее.
Рецепторная функция клеточных мембран хорошо изучена на основе действия гормона инсулина. При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
Рецепторная функция цитоплазматической мембраны также включает распознавание соседних однотипных клеток. Такие клетки прикрепляются друг к другу различными межклеточными контактами.
В тканях с помощью межклеточных контактов клетки могут обмениваться между собой информацией с помощью специально синтезируемых низкомолекулярных веществ. Одним из примеров подобного взаимодействия является контактное торможение, когда клетки прекращают рост, получив информацию, что свободное пространство занято.
Межклеточные контакты бывают простыми (мембраны разных клеток прилегают друг к другу), замковыми (впячивания мембраны одной клетки в другую), десмосомы (когда мембраны соединены пучками поперечных волокон, проникающих в цитоплазму). Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) — синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
Внеклеточная и внутриклеточная жидкости представляют собой водно-солевые растворы, в которых множество растворимых молекул диссоциируют на положительно и отрицательно заряженные атомы,— ионы. Ионы и молекулы водных растворов находятся в состоянии постоянного возбуждения и осуществляют процесс диффузии—движения из областей высокой концентрации в область меньшей концентрации. Диффузия ионов происходит не только по градиенту концентрации, но и под влиянием электрических градиентов. Положительно заряженные ионы, например ионы натрия (Na + ) и калия (К + ), называют катионами, поскольку в электрическом поле они движутся к катоду.
Отрицательно заряженные ионы, например ионы хлора (Cl - ), называют анионами, так как они движутся к аноду. Одноименные заряды (например, Na + и К + ) отталкиваются, а разноименные (например, Na + и Cl - ) — притягиваются.
Клеточную мембрану можно рассматривать в качестве электрического конденсатора, поскольку она состоит из двух противоположно заряженных слоев, между которыми расположен изолирующий липидный слой. Электрический потенциал тканевой жидкости на плазматической мембране стремится к нулю (0 мВ) за счет нейтрализующих влияний анионов Cl - на катионы Na + (и другие катионы), а в цитозоле — за счет влияний белков-анионов на катионы К + .
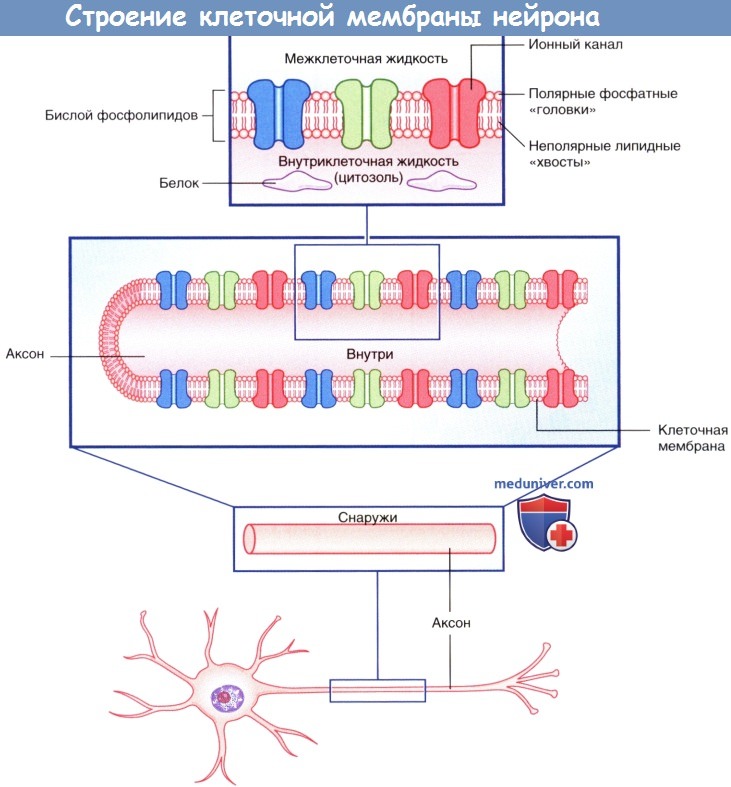
Строение клеточной мембраны нейрона.
Изображены мембранные белки, образующие ионные каналы.
а) Ионные каналы. Ионные каналы представляют собой трансмембранные белки, в центре которых находятся отверстия, по которым осуществляется транспорт ионов через клеточную мембрану. Большинство ионных каналов селективно пропускает определенные ионы, например, Na + , К + , Cl - . Выделяют несколько видов ионных каналов, из которых наиболее значимы первые три типа из следующих:
• Неуправляемые (независимые) ионные каналы постоянно находятся в открытом состоянии и осуществляют трансмембранный перенос ионов, создавая потенциал покоя на мембранах нейронов.
• Потенциалозависимые (потенциал-управляемые) ионные каналы содержат потенциалчувствительную цепь аминокислот, регулирующую открытие и закрытие отверстий ионных каналов в зависимости от изменений мембранного потенциала. Потенциал-управляемые ионные каналы играют важную роль в формировании потенциала действия.
• Энергозависимые транспортеры — ионные обменники (насосы) — обеспечивают постоянство концентраций ионов. Натрий-калиевый насос обеспечивает поддержание потенциала покоя.
• Хемоуправляемые (медиаторозависимые) ионные каналы нервной системы обеспечивают временное преобразование мембранного потенциала. В основном эти каналы представлены на постсинаптических мембранах. Активация хемоуправляемых ионных каналов может осуществляться молекулами медиаторов напрямую или опосредованно.
• Механочувствительные каналы активируются под действием физических раздражителей, вызывая деполяризацию мембраны и образование потенциалов действия, что обеспечивает восприятие определенных сигналов нервной системой. Каждый рецептор осуществляет преобразование специфических сигналов, например изменения длины или сократимости мышечных волокон, температурной и тактильной чувствительности кожи, хемочувствительности полости носа и рта или электромагнитных импульсов сетчатки.
На рисунке ниже изображены три неуправляемых ионных канала, обеспечивающих развитие потенциала покоя.
Если бы пассивная диффузия различных ионов через соответствующие им каналы осуществлялась свободно в одинаковом объеме, то не существовало бы разницы зарядов внутри и снаружи мембраны. В действительности проницаемость плазматической мембраны для ионов К + и Cl - намного выше, чем для ионов Na + , поскольку количество ионных каналов различается: калиевые каналы наиболее многочисленны, количество хлорных каналов вдвое меньше, а натриевые каналы встречают реже всего.
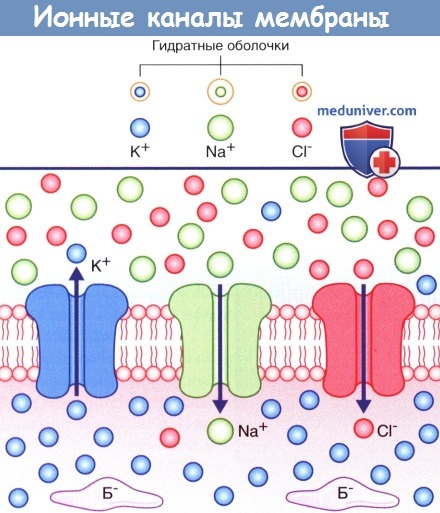
В состоянии покоя ионы Na + и Cl - сконцентрированы на внешней стороне плазматической мембраны за счет взаимного притяжения и образования толстой гидратной оболочки вокруг ионов Na + .
Ионы К + сконцентрированы на внутренней стороне плазматической мембраны за счет притяжения к белкам-анионам (Б - ).
Стрелками показано направление ионных градиентов концентрации.
б) Мембранный потенциал покоя. Мембранный потенциал невозбужденного нейрона (находящегося в состоянии покоя) формируется за счет разницы концентраций ионов Na + и К + во внеклеточной жидкости и цитозоле. Концентрация ионов К + в цитозоле выше, чем во внеклеточной жидкости в 20 раз, в то время как концентрация ионов Na + и Cl - во внеклеточной жидкости выше, чем в цитозоле в 10 и 3,8 раз, соответственно. Это обусловлено тем, что ионы К + под действием химических движущих сил переносятся из клетки в межклеточную жидкость, а ионы Na+ и Cl - — в обратном направлении.
На рисунке ниже показан вольтметр, электроды которого погружены в окружающую аксон внеклеточную жидкость. Один из электродов находится в стеклянной пипетке с тонким кончиком. В левой части рисунка, где оба электрода погружены во внеклеточную жидкость, разница электрического напряжения отсутствует, и вольтметр регистрирует нулевой показатель. В правой части рисунка пипетка проникает через плазматическую мембрану аксона в цитозоль, что позволяет оценить электрический заряд внутриклеточной жидкости. В данном случае разность электрических потенциалов составляет -70 мВ, однако у разных нейронов она может варьировать от -60 до -80 мВ.
Этот показатель отражает мембранный потенциал покоя, т.е. потенциал мембраны аксона, который не проводит электрические импульсы.
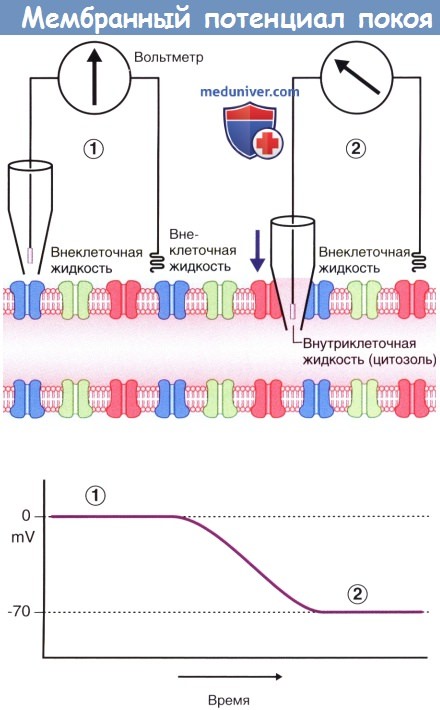
Мембранный потенциал покоя.
(1) Оба электрода вольтметра погружены во внеклеточную жидкость, окружающую аксон. Конец левого электрода находится в стеклянной пипетке.
Разница электрических потенциалов не регистрируется; на графике отмечены нулевые показатели.
(2) При опускании пипетки (направление указано стрелкой) производят пункцию плазматической мембраны для забора внутриклеточной жидкости.
Регистрируется разница электрических потенциалов величиной -70 мВ.
в) Проницаемость мембраны в состоянии покоя: 1. Ионы К + . На уровне ионного канала перемещение ионов К + осуществляется под электрическими влияниями притягивающего действия белков-анионов на внутренней стороне мембраны и отталкивающего действия ионов Na + на ее наружной стороне; при отсутствии этих факторов концентрация ионов К + на разных сторонах плазматической мембраны была бы одинаковой.
Концентрационный градиент на внешней стороне плазматической мембраны и электрический градиент потенциала на внутренней ее стороне уравновешиваются, когда мембранный потенциал достигает определенной величины, которую называют равновесным потенциалом для ионов К + (Ek). Его величина может быть рассчитана из уравнения Нернста, которое позволяет представить электрический градиент потенциала иона через его концентрационный градиент на основании законов термодинамики:
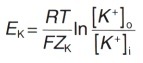
где Ek — равновесный потенциал для ионов К + (мВ)
R — газовая постоянная (8,31 Дж/(моль/°К)
Т — абсолютная температура в градусах Кельвина (310 К = 37 °С) F — число Фарадея (96500 Кл/моль)
Zk — валентность ионов К + (+1)
ln — натуральный логарифм
[К + ]0 — концентрация ионов K + с наружной стороны мембраны
[К + ]i — концентрация ионов К + с внутренней стороны мембраны.
Переведем натуральный логарифм в десятичный и рассчитаем значение показателя дроби:
Рассчитаем эти же значения для ионов Na + и Cl - :

Для расчета мембранного потенциала покоя применяют уравнение Гольдмана, учитывающее проницаемость мембраны для трех основных ионов и их приблизительное количество.
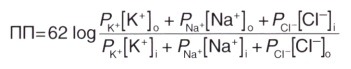
где ПП — потенциал покоя
62 — RT/Fx2,3 (постоянная для перевода натурального логарифма в десятичный)
Р — константа проницаемости мембраны для каждого из ионов (соответствует приблизительному количеству ионных каналов) о, i—концентрации ионов на внешней и внутренней сторонах мембраны, соответственно; концентрация отрицательно заряженных ионов Cl - внутри клетки расположена в числителе, а снаружи — в знаменателе дроби, поскольку log (X/Y) = log (Y/X).
Квадратными скобками обозначены концентрации ионов.
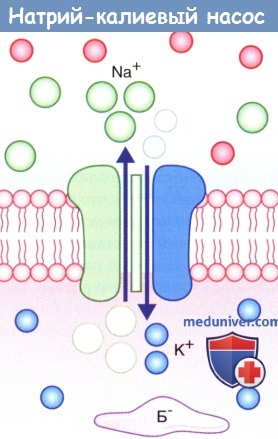
Натрий-калиевый насос.
На рисунке показан одновременный перенос трех ионов Na + из клетки в обмен на два иона К + в клетку.
Б - — белок-анион.
2. Натрий-калиевый насос. Под влиянием градиента концентрации происходит непрерывное перемещение ионов Na + внутрь клетки, а ионов К + — наружу, что нарушает постоянство потенциала покоя. Поддержание этого равновесия обеспечивает натрий-калиевый насос, корректирующий пассивное движение ионов. Натрий-калиевый насос представляет собой канал, способный перемещать ионы Na + наружу, а ионы К + — внутрь клетки. Во время работы насоса происходит перенос трех ионов Na + из клетки на каждые два иона К + в клетку.
Перенос обоих катионов осуществляется против градиента концентрации за счет энергии превращения АТФ в АДФ ферментом АТФазой. Активация этого фермента происходит при повышении концентрации ионов Na + в цитозоле.
Аксональная дегенерация при рассеянном склерозе возникает вследствие нарушения работы натрий-калиевого насоса в демиелинизированной аксолемме, что вызывает увеличение концентрации ионов Na + , которое в свою очередь приводит к повышению высвобождения ионов Са 2+ из внутриклеточного пула.
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
Каждый нейрон как структурная единица нервной ткани являет собой клетку.
Три особенности нервных клеток:
· Относительно большие размеры нервных клеток: даже у самых мелких из них линейные размеры измеряются в миллиметрах.
· Мультиполярность и неправильная форма.
· Локальные различия во внутренней структуре.
Каждый нейрон имеет клеточную или плазматичекую мембрану – плазмалемму, - определяющие границы индивидуальной клетки. С помощью мембраны и заключённых в ней молекулярных механизмов осуществляется взаимодействие нейрона с другими нейронами или улавливание изменений в локальной среде.
Клеточная мембрана состоит из двух слоёв липидных молекул. Липидные слои клеточных мембран во всех клетках одинаковы, а специфичность определяется мембранными белками. Мембранные белки – это ключ к пониманию функций нейрона, а следовательно и функций мозга, так как они изменяют натрий-калевые соотношения и этим обеспечивают проведение нервного импульса. Всё то, что находится внутри плазматической мембраны, исключая ядро, называется цитоплазмой. Цитоплазма состоит из основного вещества, включений и органелл. Органеллы – постоянные структуры цитоплазмы, выполняющие в клетке жизненно-важные функции.
Митохондрииобеспечивают клетку энергией. Особенно их много в местах отхождения аксона, в области перехватов Ранвье и синапса. В нейроне митохондрии осуществляют интенсивный энергетический обмен. Для митохондрии нервной клетки характерен короткий жизненный цикл, что связано с интенсивными процессами энергетического обмена. Характерной особенностью нейронов является наличие нейрофибрилл, образующих в теле нейрона густую сеть. Они состоят из микротрубочек, то есть тонких опорных структур, помогающих нейрону сохранять определённую форму и из нейрофиламентов, участвующих в сокращениях. Эти компоненты имеются и в других клетках, но для них характерна более упорядоченная структура.
Нейрофибриллы имеют белковую структуру. Полагают, что они участвуют в транспорте ионов и метаболитов, а также выполняют функции опоры и сокращения. Эндоплазматический ретикулум бывает гладкий (негранулярный) и шероховатый (гранулярный). Мембраны шероховатого ретикулума усеяны рибосомами, необходимых клетке для синтеза секретируемых ею белковых веществ. В области шероховатой сети происходит образование белков и липидов цитоплазматических мембран, а также их сборка.
В цитоплазме нейрона имеется обилие элементов шероховатого ретикулума, следовательно, нейроны – это клетки с весьма интенсивной секреторной деятельностью.
В нейронах имеются отдельные свободные рибосомы, которые собираются в полисомы или розетки. Рибосомы отсутствуют в аксонах.
Вещество Ниссля или тигроидное вещество
В нервных клетках гранулярная эндоплазматическая сеть, то есть система канальцев с рибосомами, образует хромофильную структуру, которая при световом микроскопе выявляется как тигроидное вещество или вещество Ниссля. Немецкий гистолог Ф. Ниссль, будучи студентом Мюнхенского университета, в 1884 году предложил и использовал митиленовый синий для окрашивания структур нервной ткани, что фактически ознаменовало начало новой эры нейроанатомии и нейропатологии. Вещество Ниссля располагается исключительно в перикарионе и начальных отделах дендритов, то есть в теле нейрона. Это наиболее специфический органоид нервной клетки, в котором осуществляется интенсивный синтез белков. Это гранулярная эндоплазматическая система – система канальцев с рибосомами. Здесь происходит, во-первых, интенсивный синтез белков, необходимых для жизнедеятельности нейрона и, во-вторых, синтез ферментов, поддерживающих нейронные градиенты.
Вещество Ниссля имеет самую разнообразную форму: так, например, в крупных двигательных нейронах это крупные многоугольные глыбки, заполняющие протоплазму, а в мелких чувствительных нейронах – густомелкозернистые глыбки.
Вещество Ниссля отсутствует в аксонах.
Электронно-микроскопическими исследованиями выявлено тонкое строение аппарата Гольджи. Этот сложный трёхмерный единый комплекс чашеобразной формы представлен собранными вместе диктиосомами (от греческого диктио – сеть). Диктиосома состоит из одной или нескольких стопок из трёх-десяти параллельных плотно упакованных уплощенных и слегка изогнутых мешочков, то есть цистерн, которые разделены тонкой прослойкой гиалоплазмы. Эти пространства не сообщаются друг с другом. Обычно к проксимальной части диктиосомы примыкают элементы эндоплазматической сети, а от дистальной её части отделяются секреторные гранулы. Элементы аппарата Гольджи располагаются около ядра вблизи клеточного центра и часто связаны с вакуолями, что особенно характерно для секретирующих клеток. В аппарате Гольджи накапливаются вещества, которые синтезируются в эндоплазматической сети. Здесь они гранулируются и в таком состоянии разносятся по клетке.
Мембраны гладкой эндоплазматической сети лишены полисом. Эта сеть функционально связана с обменом углеводов, жиров и других веществ небелковой природы.
В зоне диктиосомы различают проксимальный участок (или формирующийся цис-участок, включающий цистерны, обращённые к расширенным элементам гранулярной эндоплазматической сети) и небольшие транспортные пузырьки; а также дистальный (или зрелый транс-участок, образованный цистернами, обращёнными к вакуолям и секреторным гранулам).
Между цис- и транс-участками находится промежуточный участок, включающий небольшое количество цистерн. К дистальному участку диктиосомы - последней краевой цистерне прилегает так называемая транс-сеть Гольджи, состоящая из трубчатых элементов и множества мелких вакуолей. Она участвует в образовании лизосом, а также в разделении и сортировке белков для транспортных пузырьков.
Важнейшей функцией аппарата Гольджи является участие его мембранных элементов (то есть цистерн и пузырьков) в секреции и накоплении продуктов, синтезированных в эндоплазматической сети, а также в модификации (то есть химической перестройке белков), поступающих из гранулярного эндоплазматического ретикулума.
В цистернах аппарата Гольджи происходит синтез и сортировка модифицированных белков, а также упаковка секретированных продуктов в гранулы. Элементы аппарата Гольджи принимают участие в образовании лизосом, формировании клеточных мембран, в процессах выведения готовых секреторных продуктов за пределы клетки, к другим клеточным органеллам или плазматической мембране. Таким образом, в аппарате Гольджи происходит не просто перенос продуктов из одной полости в другую, но и постепенное их созревание и модификация белков, заканчивающееся сортировкой продуктов, направляющихся в лизосомы, к плазматической мембране или к секреторным вакуолям.
Следующая органелла – лизосома, которые образуются в аппарате Гольджи. Они впервые были открыты только в 1955 году. Это округлые пузырьки, окружённые мембраной. Лизосомы бывают разные по размерам и плотности. Они содержат большое количество ферментов (более 50), их функция – внутриклеточное переваривание различных химических соединений и структур. Они содержат гидролитические, то есть разрушающие ферменты и представляют защитно-литический аппарат нейрона. В зависимости от количества ферментов лизосома имеет различное окрашивание (меланин – чёрный, липофусцин – жёлтый, зелёный, серый).
Ядро – самая большая органелла, крупная, светло окрашенная, в центре клетки. В ядре находится хроматин, который является интерфазной формой существования хромосом. Хроматин в ядре находится в дисперсном состоянии и не образует хромосом, так как нервная клетка после рождения не делится. То есть ядро находится в интерфазе, а генетически-обусловленные продукты обеспечивают сохранение и изменение его функций на протяжении всей жизни.
Нервные отростки
Дендриты
От греческого dendron. Они образуются в процессе дифференцировки нервных клеток, позднее – нейритов. Они содержат тела и все те же органеллы, но, что особенно важно, не имеют нейроглиальной оболочки и как правило короткие и сильно ветвящиеся. По-видимому, они служат для увеличения поверхности, воспринимающей нервный импульс. Их воспринимающая поверхность в среднем в 5-10 раз превышает поверхность нейрона. Характер ветвления дендритов отражает рецептивное поле нейрона, то есть его связи с другими нейронами. Их число, порядок их отхождения от тела и характер ветвления определяют форму нейрона. Как правило, восприятие нервного импульса участвуют не только нейриты, но и тело нейрона, но иногда тело нейрона выполняет только метаболические, то есть синтезирующие функции и не участвует в восприятии нервного импульса.
Поэтому Бодиан в 1962 году предложил выделять дендритную зону для обозначения рецептивной поверхности нейрона и перекарион, то есть околоядерное, для обозначения ядра и окружающей его цитоплазмы. У большинства нейронов поверхность перекариона входит в дендритную зону, но встречаются нейроны (например, псевдоуниполярные), у которых дендритная зона может находиться на большом расстоянии от перекариона (до 1 метра).
Если импульс идёт через третий нейрон, это тормозная функция.
Непосредственно на ветвях дендритов могут образовываться синапсы, но встречаются дендриты, на ветвях которых имеются особые выросты, называемые шипиками, необходимые для образования синапсов. Их длина составляет 2 нм, а количество увеличивается от тела к периферии.
В коре больших полушарий у корковых нейронов шипики имеют особый шипиковый аппарат.
Аксон
Это единичный отросток нейрона, достигающий в длину до полутора метров, постоянного диаметра, покрытый нейроглиальными оболочками. Аксон проводит нервный импульс от тела нервной клетки к другим нейронам и рабочим органам.
Аксон начинается в виде осевого цилиндра, то есть протоплазматического продолжения нервной клетки, ещё не покрытого оболочкой. Несколько отступив от тела клетки, его обступают оболочки, которые позднее возникают у самого аксона.
Аксон покрыт двумя слоями нейроглиальной оболочки.
Непосредственно к аксону прилегает внутренний слой – миелиновая оболочка. Они появляется рядом с осевым цилиндром в виде небольших жировых капель, сливающихся в сплошную оболочку. Получая миелиновую оболочку, аксон становится основой нервного волокна.
Миелиновая оболочка выполняет несколько важнейших функций:
По-видимому, она служит изолятором нервного волокна. Жироподобное вещество миелин является электрическим изолятором. Он придаёт клеткам белую окраску, что позволило разделить всё вещество нервной системы на белое и серое. Химический состав этого липидно-белкового комплекса сложный. Миелин состоит, преимущественно, из основного липидного материала – холестерола. После липидов, то есть жировых молекул, содержащих фосфор, церебразида, то есть сложной жировой молекулы, содержащей сахар, идёт белок.
Липиды оказывают существенное влияние на конфирмационные характеристики белков. Миелин участвует в питании нервного волокна и выполняет структурную и питательную функцию. Клетки миелиновой оболочки поддерживают целостность аксона. Кроме того, она увеличивает скорость проведения нервного импульса по нервному волокну. Процесс распространения раздражения в нервной системе называется нервным импульсом. Реагирование на импульс называется нервной возбудимостью. Миелиновые волокна проводят нервный импульс значительно быстрее, чем волокна такого же диаметра, лишённые оболочки.
Немецкий учёный Герман Берингольц (автор фундаментальных трудов по физике, биофизике, физиологии и психологии) в 1852 году впервые измерил скорость распространения нервного импульса по нервному волокну. В тонких волокнах скорость проведения импульсов – не более 2 метров в секунду, тогда как в толстых миелинизированных волокнах она достигает 100 метров в секунду и более. Поэтому миелиновая оболочка поддерживается в цельном виде другой оболочкой – неврелеммой или швановской оболочкой, которая в виде тоненькой линии очерчивает контуры миелиновой оболочки.
Неврилемма представляет собой тонкий соединительно-тканный футляр, под которым располагается небольшие участки цитоплазмы с ядрами нейроглиальных клеток. Местами неврилемма прерывается, непосредственно примыкая к осевому цилиндрику, образуя перехваты Ранвье. Они разбивают миелиновый футляр осевого цилиндра на отдельные межузловые сегменты, повторяющиеся через равные промежутки, при этом каждому сегменту соответствует одна швановская клетка. В области перехватов Ранвье будут образовываться синапсы.
Полагают, что оболочки возникают вокруг аксона к тому времени, когда нерв начинает проводить импульс. А эволюционный смысл в появлении оболочки состоит в экономии метаболической энергии мозга. Нейриты образуют белое вещество головного и спинного мозга, периферические нервы и проводящие пути ЦНС
В месте отхождения аксона от тела имеется аксонный холмик.
В холмике отсутствует тигроидное вещество. Клеточная мембрана аксона называется аксолеммой, а цитоплазма – аксоплазмой.
Аксолемма выполняет важнейшую роль в проведении нервного импульса. В аксоплазме находятся нейрофибриллы, митохондрии и агранулярная эндоплазматическая сеть. Все эти органеллы сильно вытягиваются в длину.
В аксоплазме происходит постоянный ток молекул от тела нейрона к периферии и в обратном направлении.
Аксоны делятся на несколько крупных ветвей, которые отходят от перехватов Ранвье. Эти ветви оканчиваются конечными ответвлениями, которые называется терминалиями, которые, в свою очередь, образуют синапсы от других нейронов и рабочих органов.
Аксон всегда покрыт нейроглиальной оболочкой. В зависимости от характера её структуры различают два типа волокон: немиелинизированные, то есть безмякотные, и миелинизированные или мякотные волокна.
Первый тип волокон, то есть немиелинизированные встречается главным образом в вегетативной нервной системе и имеет малый диаметр. Такой аксон погружён в нейроглиальную клетку так, что оболочка нейроглиальной клетки охватывает его со всех сторон, образуя мезаксон.
Установлено, что в одну нейроглиальную клетку может погружаться до 10-20 аксонов. Такие волокна называются волокнами кабельного типа. При этом оболочка образует цепочка нейроглиальных клеток.
Немиелинизированные аксоны имеют меньший диаметр
Миелиновая оболочка
Протяжённость миелиновой оболочки начинается несколько отступив от начала аксона и заканчивается двух микрон от синапса. Она состоит из отдельных цилиндров равной длины 1,5-2 микрона, которые называются межузловыми сегментами, разделёнными перехватами Ранвье.
В области перехватов аксон либо обнажён, либо покрыт неврилеммой (в периферической нервной системе). Также там могут отходить ветви и образовываться синапсы.
Миелиновая оболочка – упорядоченная протеидная структура, состоящая из чередующихся белковых и липидных слоёв. Её структурной единицей является бимолекулярный липидный слой, заключённый между двумя мономолекулярными белковыми слоями, причём количество слоёв достигает 100 микрон.
Оболочка является изолятором и обладает большим сопротивлением постоянному току, что способствует огромному ускорению в проведении нервного импульса. Нервный импульс здесь перескакивает с одного перехвата Ранвье на другой, так как деполяризация аксона происходит только в области перехватов Ранвье.
Такое проведение нервного импульса называется скачкообразным или сальтоторным.
Процесс миелинизации
В периферической нервной системе миелиновая оболочка образуется в результате спирального накручивания вокруг аксона мезаксона нейроглиальной клетки. При этом число витков нарастает по мере роста аксона.
Следовательно, субъединицей миелиновой оболочки является участок клеточной мембраны швановской клетки. Цитоплазма и ядро её оттесняются на периферию, образуя неврилемму, которая также называется швановской клеткой.
В центральной нервной системепроцесс миелинизации менее упорядоченный. Оболочка здесь образуется в результате спирального накручивания вокруг аксона отростка олигодендроцита, причём отростки одного олигодендроцита накручиваются вокруг нескольких аксонов.
В периферической нервной системе в миелиновой оболочке образованы насечки Шмидта-Лантермана, то есть косо расположенные воронкообразные щели. Полагают, что они соединяют цитоплазму нейроглиальной клетки, располагаясь снаружи и внутри миелиновой оболочки.
Синапс
Нейрон может образовывать от ста до тысячи синапсов и сам получать информацию от 1000 других нейронов.
Преобладают синапсы аксодендритные (аксон – дендриты нейрона) и аксосоматические (аксон – тело нейрона). Встречаются также аксоаксонные или аксоаксональные синапсы. Им приписывают тормозную функцию, так как они находятся там, где один из аксонов синаптирует с дендритом третьего нейрона. Реже встречаются соматосоматические, дендродендритные и соматодендритные синапсы. У животных и человека преобладают синапсы химические, так как передача нервных импульсов осуществляется не ими, а через посредство особых химических веществ – медиаторов.
В нервной системе известно около 30 медиаторов. Наиболее известные – ацетилохолин и катехоламины (норадреналин и другие). Это химические вещества, молекулы которых имеют небольшую длину с положительно-заряженным атомом азота. Среди медиаторов имеются также нейропептиды, то есть короткие цепочки аминокислот. Медиаторы синтезируются в нервных окончаниях.

В области синапса нервные окончания теряют миелиновую оболочку и образуют расширение, которое называется синаптической мембраной диаметром примерно 1 микрон, покрытую пресинаптической мембраной. Между ними находится синаптическая щель. Синаптические пузырьки, круглые или овальные, содержащих по 10-100 тысяч молекул медиатора. Молекулы медиатора соединяются с рецепторными пузырьками постсинаптических мембран, что вызывает образование постсинаптического потенциала. Затем сразу молекулы медиатора инактивируются, то есть разрушаются или возвращаются в синаптические пузырьки.
Структура синапса такова, что нервный импульс может идти только в одном направлении, то есть, иными словами, синапс поляризован, что и определяет односторонность проведения нервного импульса по аксону.
Нейроглия
Нейроглиальные клетки выполняют ряд функций: опорную, трофическую, секреторную, разграничительную и защитную. Существует макроглия и микроглия.
Клетки макроглии развиваются из общей закладки с нейронами, то есть из эктодермы, но, в отличие от нейронов, делятся в течение всей жизни, имеют отростки только одного типа и не образуют синапсов.
Клетки микроглии имеют мезодермальное происхождение и проникают в нервную ткань вскоре после рождения.
Читайте также:
