
Ядра спинномозговых и черепных нервов как сегментарные центры
ВОПРОСЫ ПРАКТИЧЕСКОГО КУРСА
- Спинной мозг: топография, внешнее и внутреннее строение. Скелетотопия сегментов спинного мозга.
- Топография и состав белого и серого вещества спинного мозга.
- Анатомия и внутренний состав корешков спинного мозга. Конский хвост.
- Анатомия спинномозгового нерва, его формирование и ветви.
- Топография, компоненты и внешнее строение ствола головного мозга (продолговатого мозга, моста, среднего мозга).
- Топография и элементы внешнего строения мозжечка. Ядра мозжечка. Ножки мозжечка.
- Стенки и сообщения 4-го желудочка и водопровода мозга. Анатомия ромбовидной ямки. Сосудистая основа и сосудистое сплетение 4-го желудочка.
- Топография и природа ядер черепных нервов.
- Чувствительные ядра черепных нервов: нейронный состав, топография, принципиальные связи, виды и зоны иннервации.
- Двигательные ядра черепных нервов: нейронный состав, топография, принципиальные связи, зоны иннервации.
- Автономные (вегетативные) ядра черепных нервов: нейронный состав, топография, принципиальные связи, виды и зоны иннервации.
- Топография надсегментарных центров ствола головного мозга (ядра оливы, ретикулярной формации, черного вещества, красного ядра, ядер 4-холмия). Собственные ядра моста как релейные центры в связях новой коры и нового мозжечка.
- Компоненты белого вещества ствола головного мозга и их природа (мозговые полоски, трапециевидное тело, петли - спинномозговая, медиальная, латеральная, тройничная; медиальный продольный пучок, ручки холмиков, пирамидные и экстрапирамидные пути).
- Анатомия и краткая функциональная характеристика компонентов таламической области (таламического мозга).
- Анатомия и краткая функциональная характеристика компонентов гипоталамуса.
- Стенки и сообщения 3-го желудочка. Его сосудистая основа и сосудистое сплетение.
- Топография и классификация базальных ядер конечного мозга.
- Внутренняя капсула, ее отделы. Топография составляющих ее проводящих путей.
- Лимбическая доля. Анатомия центрального и периферического отделов обонятельного мозга.
- Борозды и извилины поверхностей полушарий большого мозга.
- Локализация корковых анализаторов 1-й и 2-й сигнальных систем.
- Стенки и сообщения боковых желудочков головного мозга.
- Организация проводящих путей кожной чувствительности.
- Организация проприоцептивных проводящих путей мозжечкового и коркового направлений.
- Организация пирамидных путей.
- Организация старых и нового экстрапирамидных путей.
- Анатомия комиссуральных связей головного мозга (мозолистое тело, передняя спайка, спайка свода).
- Оболочки спинного мозга и их производные. Межоболочечные пространства и их содержимое.
- Оболочки головного мозга и их производные. Подпаутинное пространство и его части, содержимое.
- Пути циркуляции спинномозговой жидкости.
- Рентгеноанатомия желудочков мозга.
Сегментарный аппарат ствола головного мозга представляет собой совокупность анатомически и функционально взаимосвязанных структур, предназначенных для осуществления безусловных (врожденных) рефлексов, замыкающихся на уровне ствола головного мозга. Примерами таких рефлексов являются сосательный, глотательный, роговичный, кашлевой и т.д.
В состав сегментарного аппарата ствола головного мозга входят следующие структуры.
- 1. Корешковые волокна черепных нервов, включающих чувствительный компонент, – V пара (тройничный нерв), VII пара (лицевой нерв), IX пара (языкоглоточный нерв), X пара (блуждающий нерв). Они представляют собой расположенные в веществе ствола головного мозга центральные отростки псевдоуниполяриых клеток тройничного узла (V пары), коленчатого узла (VII пары), верхнего и нижнего узлов (IX и X пар). Корешковые волокна заканчиваются синаптическими окончаниями на вставочных нейронах ствола головного мозга.
- 2. Вставочные нейроны, роль которых выполняют рассеянные клетки ретикулярной формации ствола головного мозга. Аксоны этих клеток синаптически заканчиваются на нейронах двигательных ядер черепных нервов.
- 3. Мультиполярные нейроны двигательных ядер черепных нервов – III пара (глазодвигательный нерв), IV пара (блоковый нерв), V пара (тройничный нерв), VI пара (отводящий нерв), VII пара (лицевой нерв), IX пара (языкоглоточный нерв), X пара (блуждающий нерв), XI пара (добавочный нерв) и XII пара (подъязычный нерв).
- 4. Часть аксонов нейронов двигательных ядер черепных нервов, составляющих двигательные корешковые волокна в пределах вещества мозга.
Остальные элементы рефлекторных дуг безусловных рефлексов относят к периферической нервной системе (корешковые волокна, лежащие за пределами ствола головного мозга, краниальные чувствительные ганглии, черепные нервы и их ветви).
В большинстве случаев вставочные нейроны сегментарного аппарата ствола головного мозга обеспечивают передачу нервных импульсов на нейроны двигательных ядер нескольких черепных нервов, причем не только своей, но и противоположной стороны. Например, при раздражении кожи лица в области щеки или губ у новорожденного возникают сосательные движения. Воспринимают раздражение рецепторы, являющиеся окончаниями псевдоуниполярных клеток узла тройничного нерва. Распространение нервного импульса в стволе головного мозга осуществляется на нейроны двигательных ядер V, VII, IX, X, XI и XII пар черепных нервов. В связи с этим в осуществлении сосательного акта принимают участие жевательные, мимические мышцы, мышцы неба, глотки, шеи и языка. При этом мускулатура включается в осуществление ответной реакции в равной степени как на своей, так и на противоположной стороне тела.
Ретикулярная формация – это комплекс анатомически и функционально взаимосвязанных нейронов шейного отдела спинного мозга и ствола головного мозга, окруженных множеством волокон, идущих в различных направлениях. Нейроны ретикулярной формации имеют длинные маловетвящиеся дендриты и аксон, отдающий значительное количество вторичных ветвей. Это позволяет нейрону установить контакты примерно с 25 тыс. других нейронов. Ветви нейронов под микроскопом образуют своеобразную сеть – ретикулум. Именно сетевидное расположение волокон, связывающих между собой нервные клетки, и послужило основой для предложенного названия.
Структурные элементы ретикулярной формации в шейных и верхнегрудных сегментах спинного мозга локализуются между задним и боковым рогами, в ромбовидном и среднем мозге – в покрышке, в промежуточном мозге – в составе таламуса.
Наряду с многочисленными отдельно лежащими нейронами, различными по форме и величине, в стволе головного мозга имеются ядра ретикулярной формации. Рассеянные нейроны ретикулярной формации прежде всего играют важную роль в обеспечении сегментарных рефлексов, замыкающихся на уровне ствола головного мозга. Они выступают в качестве вставочных нейронов при осуществлении таких рефлекторных актов, как глотание, роговичный рефлекс и т.д.
Выяснено значение и многих ядер ретикулярной формации. Так, ядра, расположенные в продолговатом мозге, имеют связи с вегетативными ядрами блуждающего и языкоглоточного нервов, симпатическими ядрами спинного мозга. Поэтому они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и т.д.
Ядра Кахаля и Даркшевича, относящиеся к ретикулярной формации среднего мозга, посредством медиального продольного пучка имеют связи с ядрами III, IV, VI, VIII и XI пар черепных нервов. Они координируют работу этих нервных центров, что очень важно для обеспечения сочетанного поворота головы и глаз. Ретикулярная формация ствола головного мозга имеет большое значение в поддержании тонуса скелетной мускулатуры, посылая тонические импульсы на гамма-мотонейроны двигательных ядер черепных нервов и двигательных ядер передних рогов спинного мозга.
В процессе эволюции из ретикулярной формации выделились такие самостоятельные образования, как красное ядро, черное вещество. Кроме того, ряд ядер ретикулярной формации ствола головного мозга в процессе эволюции приобрели роль жизненноважных центров: дыхательный и сосудодвигательный центры продолговатого мозга; располагающиеся в промежуточном мозге центры терморегуляции, голода и насыщения, вегетативных функций и т.д.
К рассеянным клеткам и ядрам ретикулярной формации подходят коллатерали от спинномозговой, медиальной, тройничной и латеральной петель или непосредственно от чувствительных ядер черепных нервов. От нейронов ретикулярной формации эфферентные волокна направляются к двигательным ядрам черепных нервов, к мозжечку, к двигательным ядрам передних рогов спинного мозга.
Основным нисходящим трактом является ретикулярноспинномозговой путь, который берет начало в стволе головного мозга и идет к нейронам передних рогов спинного мозга, отдавая коллатерали к двигательным ядрам черепных нервов. Этот путь проводит тонические импульсы к указанным образованиям.
От нейронов медиальных и ретикулярных ядер зрительного бугра к различным областям коры полушарий большого мозга идут таламо-корковые волокна. Особенностью этих путей является диффузный характер их распределения – они заканчиваются не только во всех областях, но и во всех слоях коры полушарий большого мозга. В связи с этим в кору поступают неспецифические афферентные импульсы из ретикулярной формации спинного мозга и ствола головного мозга. Неспецифические афферентные импульсы осуществляют активацию коры полушарий большого мозга, необходимую для восприятия специфических раздражений. Последние поступают в проекционные центры коры по специализированным афферентным путям от коммуникационных ядер таламуса и коленчатых тел. Следует подчеркнуть важную роль неспецифических афферентных ретикулярных волокон в отборе информации (дифференцированном проведении импульсов), поступающей к коре полушарий большого мозга. Прерывание потока неспецифических афферентных импульсов приводит к снижению тонуса коры, апатии и наступлению сна.
Необходимо отметить, что кора полушарий большого мозга в свою очередь посылает по корково-ретикулярным путям импульсы в ретикулярную формацию. Эти импульсы возникают в основном в коре лобной доли и проходят в составе пирамидных путей. Корково-ретикулярные связи оказывают либо тормозное, либо возбуждающее действие на ретикулярную формацию ствола головного мозга, осуществляют корректировку прохождения импульсов по эфферентным путям (отбор эфферентной информации).
Таким образом, между ретикулярной формацией и корой полушарий большого мозга имеется двусторонняя связь, которая обеспечивает саморегуляцию в деятельности нервной системы. От функционального состояния ретикулярной формации зависят тонус мускулатуры, работа внутренних органов, настроение, концентрация внимания, память и т.д.
В целом ретикулярная формация создает и поддерживает условия осуществления сложной рефлекторной деятельности с участием коры полушарий большого мозга, выполняя следующие основные функции:
- 1) обеспечение сегментарных рефлексов – рассеянные клетки выступают в качестве вставочных нейронов (глотание, чихание, роговичный рефлекс, зрачковый рефлекс и т.д.);
- 2) поддержание тонуса скелетной мускулатуры – клетки ретикулярной формации посылают тонические импульсы на двигательные ядра черепных и спинномозговых нервов, преимущественно по ретикулярно-спинномозговому тракту;
- 3) коррекция проведения нервных импульсов – благодаря ретикулярной формации импульсы могут либо существенно усиливаться, или существенно ослабляться в зависимости от состояния нервной системы;
- 4) активное влияние на высшие центры коры полушарий, что приводит либо к снижению тонуса коры, апатии и наступлению сна, либо к повышению работоспособности, эйфории, участвуя в регуляции сна и бодрствования и т.д.;
- 5) участие в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и других вегетативных функций (центры ствола головного мозга).
Вегетативная система обеспечивает адаптацию к меняющимся факторам среды посредством: коры, диэнцефальной области, ядер среднего и продолговатого мозга, боковых рогов, узлов в симпатическом стволе.
Вегетативную нервную систему разделяют на сегментарный и надсегментарный отделы.
Сегментарный отдел вегетативной нервной системы обеспечивает вегетативную иннервацию отдельных сегментов тела и внутренних органов, которые к ним относятся. Он подразделяется на симпатическую и парасимпатическую часть.
Парасимпатическая часть вегетативной нервной системы делится на краниобульбарный и сакральный отделы. Вегетативные ядра глазодвигательного нерва - добавочное (ядро Якубовича) и центральное заднее (ядро Перлиа) находятся на уровне среднего мозга. Их аксоны в составе глазодвигательного нерва идут к ресничного узлу, который находится в заднем отделе орбиты. От него постганглионарные волокна в составе коротких цилиарных нервов иннервируют гладкие мышцы глаза: мышцу, суживающую зрачок, и ресничную мышцу, сокращение которой обеспечивает аккомодацию.
В области моста находятся секреторные слезовыделительные клетки, аксоны которых в составе лицевого нерва идут к крылонебному узлу и иннервируют слезную железу. В стволе мозга также локализуются верхнее и нижнее секреторные слюновыделительные ядра, аксоны от которых идут с языкоглоточным нервом к околоушному узлу и с промежуточным нервом к подчелюстному и подъязычному узлам и иннервируют соответствующие слюнные железы.
На уровне продолговатого мозга находится заднее (висцеральное) ядро блуждающего нерва, парасимпатические волокна которого иннервируют сердце, пищеварительный канал, желудочные железы и другие внутренние органы (кроме органов малого таза).
Сакральная (крестцовая) часть вегетативной нервной системы представлена клетками, которые размещаются в боковых рогах спинного мозга на уровне S2-S4 сегментов (боковое промежуточное ядро). Аксоны этих клеток формируют тазовые нервы которые иннервируют мочевой пузырь, прямую кишку и половые органы.
Ретикулярная формация - дыхательный и сосудовигательный центры, центры сердечной деятельности, регуляции обмена веществ и т.д.).
Мозжечок - трофические центры.
Гипоталамус - главный подкорковый центр -(уровень обмена веществ, терморегуляция).
Полосатое тело - безусловнорефлекторная регуляция вегетативных функций.
Высшим центром регуляции вегетативных и соматических функций, их координации является кора полушарий большого мозга.
Симпатические нейроны расположены в боковых рогах (С8-L2), аксоны идут с передними корешками выходят из позвоночного канала и подходят к симпатическому стволу. Симпатический ствол состоит из узлов: 3 шейных, 12 грудных, 4 поясничных, 4 тазовых; преганглионарные волокна частично прерываются в узлах, частично нет и идут к вертебральным ганглиям, далее волокна идут к вертебральным узлам или сплетениям (солнечное, чревное, сердечное, легочное).
При поражение верхнего шейного узла триада Горнера ( птоз, миоз, энофтальм)
Нижнего шейного узла – расстройство сердечной деятельности, тахикардия, повышение АД.
Нарушение мышечного тонуса, контрактуры, дрожание, изменение чувствительности по типу гиперпатии, боли, нарушение потоотделения, сосудистые трофические расстройство кожи, ПЖК и костей, боли и спазмы всех органов.
Общие принципы формирования периферических нервов
Периферический отдел нервной системы представлен комплексом образований, построенных из нервной ткани, в который включают:
· корешки периферических нервов (черепных и спинномозговых),
· собственно периферические нервы,
· ветви спинномозговых и черепных нервов
· нервные сплетения (соматические или вегетативные)
· узлы-ганглии (соматические или вегетативные)
· нервные окончания (рецепторные и эффекторные)
Рис. Схема рефлекторной дуги на уровне спинного мозга. Образование спинномозгового нерва.
Спинномозговые нервы образуются из слияния соответствующих одному сегменту пар передних и задних корешков спинного мозга. В результате образуется ствол спинномозгового нерва, который всегда является смешанным, так как задний корешок – чувствительный, а передний – двигательный и с С8 по L2 также содержит еще и вегетативные преганглионарные волокна.
Отличие черепных нервов от спинномозговых
Черепные нервы имеют ряд существенных особенностей в строении. Для того чтобы приступить к изучению черепных нервов, необходимо вспомнить ряд вопросов строения центральной нервной системы, в частности проекцию чувствительных, двигательных и вегетативных ядер черепных нервов на стволовую часть головного мозга.
Ядра – это скопление серого вещества ствола мозга (тел нервных клеток), выполняющих сходные или взаимосвязанные функции. В зависимости от этого разделяют двигательные, вегетативные (парасимпатические) и переключательные-чувствительные ядра. В зависимости от того, с какими ядрами связаны черепные нервы, они будут иметь соответствующие им волокна и выполнять определенные функции.
Рис. Ствол мозга с проекцией ядер.
Так, в области среднего мозга проецируются ядра ΙΙΙ(двигательное и парасимпатическое) и IV (двигательное) пар черепных нервов,V (чувствительное).
В мосту - ядра V (три чувствительных, одно двигательное), VI (двигательное), VII (двигательное, чувствительное, парасимпатическое), на границе моста и продолговатого мозга - VIII (2 чувствительных улитковых, 4 чувствительных преддверных) пар черепных нервов.
В продолговатом мозге - ядра IX (двигательное, чувствительное, парасимпатическое), X (двигательное, чувствительное, парасимпатическое), XI (двигательное), XII (двигательное) пар черепных нервов.
Исходя из этого, легче запомнить места выхода этих нервов из ствола мозга и функциональный состав их волокон.
Следует заметить, что Ι, ΙΙ, VIII пары черепных нервов являются частями анализаторов, и поэтому являются сугубо чувствительными нервами.
ΙV,VI,XI,XII пары являются чисто двигательными нервами. Все остальные ΙΙΙ,V,VII,IX,X пары – смешанными нервами, и имеют разные корешки. Чувствительные – являются центральными отростками чувствительных клеток, расположенных в специальных ганглиях, которые заходят в вещество мозга.Двигательные и вегетативные корешки являются аксонами клеток соответствующих ядер и выходят из вещества мозга.
Необходимо выучить названия, расположение и функциональное предназначение каждого из ядер черепных нервов. Этому будут посвящены последующие разделы учебно-методического пособия.
ПРИНЦИПИАЛЬНЫЕ ОТЛИЧИЯ ЧЕРЕПНЫХ ОТ СПИННОМОЗГОВЫХ НЕРВОв
| черепные нервы | спинномозговые нервы | |
| центры (ядра) | В стволовой части головного мозга | Ядра серого вещества спинного мозга, кроме боковых рогов C8 - L2 сегментов спинного мозга |
| состав корешков нервов | Могут быть сугубо чувствительными, или двигательными, или смешанными в зависимости от того, какие ядра имеет нерв | Задние корешки чувствительные, передние - двигательные (шейный отдел), смешанные (двигательные и вегетативные) в грудном, поясничном и крестцовом) |
| места выхода | Отверстия черепа | Межпозвонковые отверстия |
| принцип иннервации | Несегментарный, преимущественно органы головы и шеи | Сегментарный Преимущественно органы туловища и конечностей |
Дата добавления: 2015-07-07 ; просмотров: 3285 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Аксоны третьих нейронов образуют таламокорковый путь (tractus thalamocorticalis), проходящий через середину задней ножки внутренней капсулы и заканчивающийся в коре полушария большого мозга.
Корковый конец интероцептивного анализатора включает в себя нижний отдел постцентральной извилины, предцентральную извилину, премоторную зону лобной доли, а также извилины височной доли.
3. Двигательные проводящие пути
Помимо эфферентных путей в состав экстрапирамидной системы входят нервные центры и их связи. К этим нервным центрам относятся чечевицеобразное ядро, хвостатое ядро, красное ядро, черное вещество, ядра ретикулярной формации, ядро оливы и др. Ведущая роль в этой группе центров принадлежит хвостатому ядру и чечевицеобразному ядру. С центрами экстрапирамидной системы имеют тесные функциональные связи зрительный бугор, гипоталамус и мозжечок. Среди многочисленных связей центров экстрапирамидной системы следует выделить двусторонние связи с корой полушарий большого мозга, особенно с премоторной областью.
Важнейшими экстрапирамидными путями являются: красноядерно-спинномозговой путь, крышеспинномозговой путь, ретикулоспинномозговой путь, преддверно-спинномозговой путь, оливоспинномозговой путь.
Поражение экстрапирамидной системы сопровождается нарушениями движений в виде их ослабления (гипокинезия) или усиления (гиперкинезы). Нарушается и мышечный тонус в сторону повышения (ригидность) или понижения (гипотония).
Красное ядро (nucl. ruber) является главным подкорковым центром экстрапирамидной системы и расположено в покрышке среднего мозга (рис. 16).
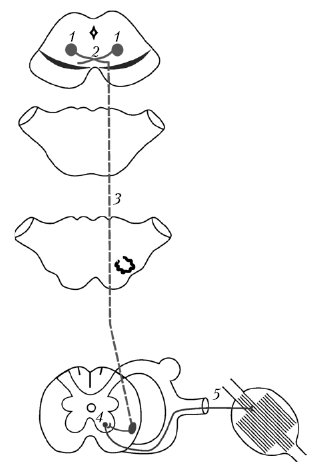
Рис. 16. Красноядерно-спинномозговой путь: 1 — красное ядро, 2 — передний (вентральный) перекрест покрышки, 3 — красноядерно-спинномозговой путь, 4 — двигательные ядра передних рогов спинного мозга, 5 — спинномозговые нервы
От крупных мультиполярных нейронов, локализованных в каудальной части красного ядра, начинается красноядерно-спинномозговой путь (tractus rubrospinalis), или пучок Монакова.
Tractus rubrospinalis истончается в каудальном направлении, так как его волокна посегментно заканчиваются в пределах ствола и спинного мозга. Этот путь располагается в покрышке среднего мозга — вентральнее и медиальнее красных ядер. В мосту он находится также в покрышке, но занимает в ней дорсолатеральное положение. Сходная локализация сохраняется и в продолговатом мозге. В спинном мозге tractus rubrospinalis расположен в боковом канатике вентральнее бокового корково-спинномозгового пути.
Гамма-мотонейроны ствола и спинного мозга, испытавшие влияние красного ядра, передают двигательные импульсы по своим аксонам в составе черепных и спинномозговых нервов к иннервируемым скелетным мышцам-сгибателям.
Следует сказать, что красное ядро само испытывает многочисленные влияния со стороны коры полушарий большого мозга, ядер стриопаллидарной системы (базальные ядра) и промежуточного мозга, а также со стороны мозжечка. Обработав всю полученную информацию, красное ядро посылает по tractus rubrospinalis такую импульсацию, которая, в конце концов, приведет к выполнению сложных привычных автоматизированных движений (ходьба, бег, трудовые и бытовые движения) или к сохранению определенной позы в течение длительного времени за счет поддержания тонуса скелетных мышц.
Этот путь считают самым филогенетически молодым среди экстрапирамидных.
Он начинается в верхних холмиках среднего мозга (другие авторы считают, что tractus tectospinalis начинается в ядрах как верхних, так и нижних холмиков), куда поступает чувствительная информация от ядра верхнего холмика (зрительная), от ядра нижнего холмика (слуховая), от ядра сосочкового тела (обонятельная) и по коллатералям спинномозговой, медиальной и тройничной петель (тактильная). Таким образом, tractus tectospinalis участвует в безусловно-рефлекторных двигательных реакциях на сильные и внезапные зрительные, слуховые, обонятельные и тактильные воздействия (для человека в этом потоке афферентных импульсов наиболее значимыми являются зрительные и слуховые).
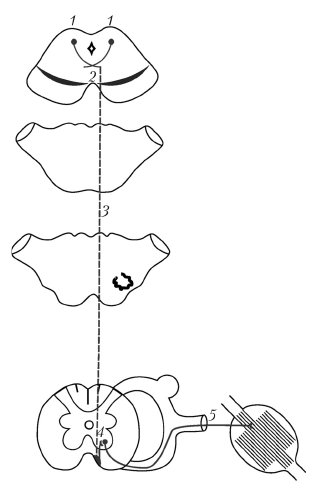
Рис. 17. Крышеспинномозговой путь: 1 — ядра верхнего холмика (пластинки четверохолмия), 2 — задний (дорсальный) перекрест покрышки, 3 — крышеспинномозговой путь, 4 — двигательные ядра передних рогов спинного мозга, 5 — спинномозговые нервы
Презентация была опубликована 3 года назад пользователемwwwwwwww wwwwww
Презентация на тему: " ЛЕКЦИЯ 6 СТВОЛ ГОЛОВНОГО МОЗГА ЯДРА ЧЕРЕПНЫХ НЕРВОВ Рис. столбы." — Транскрипт:
4 СТВОЛ ГОЛОВНОГО МОЗГА
8 ЯДРА ЧЕРЕПНЫХ НЕРВОВ Рис. столбы
9 ЯДРА НЕРВОВ КАК СЕГМЕНТАРНЫЕ ЦЕНТРЫ ЯДРО НЕРВА– группа однотипных нейронов, которыми нерв начинается либо на которых завершается ТИПЫ ЯДЕР НЕРВОВ (сегментарных центров): 1. ЧУВСТВИТЕЛЬНЫЕ ЯДРА НЕРВОВ содержат тела вставочных нейронов, которые первыми в ЦНС получают информацию с периферии через соответствующий нерв СУЩЕСТВУЮТ ТОЛЬКО В СОСТАВЕ СПИННОГО МОЗГА И СТВОЛОВОЙ ЧАСТИ ГОЛОВНОГО ПРИ ИХ ПОРАЖЕНИИ : анестезия (потеря чувствительности определенного типа в зависимости от характера ядра) в зоне действия ЭТОГО нерва
10 ЯДРА НЕРВОВ КАК СЕГМЕНТАРНЫЕ ЦЕНТРЫ ТИПЫ ЯДЕР НЕРВОВ (сегментарных центров): 2. ДВИГАТЕЛЬНЫЕ ЯДРА НЕРВОВ Содержат тела двигательных соматических нейронов, аксоны которых составляют двигательную порцию нерва Поражение двигательного ядра : периферический (вялый) паралич иннервируемых этим нервом мышц (мышечной группы): - атония - арефлексия - атрофия
11 НЕЙРОННЫЕ АНСАМБЛИ – НЕРВНЫЕ ЦЕНТРЫ НЕРВНЫЙ ЦЕНТР (морфологически, клинически) – группа однотипных нейронов с общей историей, функциями и связями и обязательным наличием синапсов СЕГМЕНТАРНЫЕ ЦЕНТРЫ ОБЛАДАЮТ НЕПОСРЕДСТВЕННОЙ, ПРЯМОЙ СВЯЗЬЮ С ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМОЙ, С ИННЕРВИРУЕМОЙ ПЕРИФЕРИЕЙ ПРЕДСТАВЛЕНЫ ЯДРАМИ СПИННОМОЗГОВЫХ И ЧЕРЕПНЫХ НЕРВОВ СУЩЕСТВУЮТ ТОЛЬКО В СОСТАВЕ СПИННОГО МОЗГА И СТВОЛОВОЙ ЧАСТИ ГОЛОВНОГО ПРИ ИХ ПОРАЖЕНИИ ПРОЯВЛЯЮТСЯ ОТЧЕТЛИВЫЕ СИМПТОМЫ В СТРОГО ОЧЕРЧЕННОЙ ОБЛАСТИ
13 Поперечный срез продолговатого мозга
17 Поперечный срез среднего мозга
19 РАСПРЕДЕЛЕНИЕ ЦЕНТРОВ В ЦНС. СЕГМЕНТАРНЫЕ - ядра нервов-ТОЛЬКО В СПИННОМ МОЗГЕ И СТВОЛЕ А ВСЁ ОСТАЛЬНОЕ .
21 НЕЙРОННЫЕ АНСАМБЛИ – НЕРВНЫЕ ЦЕНТРЫ НАДСЕГМЕНТАРНЫЕ ЦЕНТРЫ НЕ ИМЕЮТ НЕПОСРЕДСТВЕННОЙ СВЯЗИ С ИННЕРВИРУЕМОЙ ПЕРИФЕРИЕЙ (. ), ДЕЙСТВУЮТ ТОЛЬКО ЧЕРЕЗ СЕГМЕНТАРНЫЕ ЦЕНТРЫ ИЛИ ВЗАИМОДЕЙСТВУЯ ДРУГ С ДРУГОМ (. ) ПРЕДСТАВЛЕНЫ В СТВОЛЕ ГОЛОВНОГО МОЗГА И ЕГО НАДСТВОЛОВОЙ ЧАСТИ ПРИ ИХ ПОРАЖЕНИИ ОТЧЕТЛИВЫХ СИМПТОМОВ (ПАРАЛИЧЕЙ, АНЕСТЕЗИИ) нет, ВОСПРИЯТИЕ И ДВИГАТЕЛЬНЫЕ РЕАКЦИИ сохраняются, изменяется их качество (исключение – ряд центров коры полушарий большого мозга)
22 ВОТ ОСНОВНЫЕ ИЗ ЭТИХ ЦЕНТРОВ СТВОЛА: Ретикулярная формация Ядро оливы Собственные ядра моста Красное ядро Черная субстанция Центры 4-холмия
29 Archicerebellum вестибулярный: flocculus, nodulus + n. fastigii связи вестибулярные
30 Paleocerebellum Спинальный: vermis + n. globosus+ n. emboliformis проприоцептивные связи со спинным мозгом
31 Neocerebellum Neocerebellum – полушария мозжечка + n. dentatus Связи с новой корой через ядра моста
32 ПОЛОЖЕНИЕ NEOCEREBELLUM в связях NEOCORTEX
33 Вместе с мозжечком, многими надсегментарными центрами промежуточного и конечного мозга надсегментарные центры ствола ГМ составляют мощную двигательную ЭКСТРАПИПАМИДНУЮ систему, EPS – QUALITY OF ACTION обеспечивающую Вместе с мозжечком, многими надсегментарными центрами промежуточного и конечного мозга надсегментарные центры ствола ГМ составляют мощную двигательную экстрапирамидную систему, обеспечивающую качественную реализацию двигательных программ через регуляцию мышечного тонуса, координацию действия мышечных групп. ЭПС – качество действия.
35 ПРОМЕЖУТОЧНЫЙ МОЗГ - DIENCEPHALON
57 Цитоархитектонические поля Бродмана Поля Бродмана отделы коры больших полушарий головного мозга, отличающиеся по своей цитоархитектонике (строению на клеточном уровне). Выделяется 52 цитоархитектонических поля Бродмана.
59 СОМАТОТОПИЧЕСКАЯ ПРОЕКЦИЯ ЧАСТЕЙ ТЕЛА В КОРУ БОЛЬШОГО МОЗГА
61 АСИММЕТРИЯ ЛЕВОГО И ПРАВОГО ПОЛУШААРИЙ
В грудном отделе формирующегося спинного мозга с обеих сторон можно выделить четыре отдельных столба клеток серого вещества. В области базальной пластинки общий соматический эфферентный (ОСЭ) столб иннервирует поперечно-полосатую мускулатуру туловища и конечностей. Общий висцеральный эфферентный (ОВЭ) столб содержит преганглионарные нейроны вегетативной нервной системы. В области крыльной пластинки общий висцеральный афферентный (ОБА) столб получает афферентные нервные волокна от внутренних органов грудной и брюшной полостей.
Общий соматический афферентный (ОСА) столб получает афферентные волокна от конечностей и стенок полостей тела.
В стволе мозга также можно обнаружить четыре этих столба, однако они представлены отдельными фрагментами, и не каждый столб вносит свой вклад в формирование всех черепных нервов. Здесь столбы имеют следующие связи.
• ОСЭ столб. Иннервирует поперечно-полосатые мышцы глазницы (посредством глазодвигательного, блокового, отводящего нервов) и языка (посредством подъязычного нерва).
• ОВЭ столб. Дает начало парасимпатической системе черепа. Волокна следуют к ресничному, ушному, крылонебному, поднижнечелюстному ганглиям головы и шеи, а также к блуждающим ганглиям шеи, грудной клетки и брюшной полости.
• ОВА столб. Получает нервные импульсы от области иннервации языкоглоточного и блуждающего нервов.
• ОСА столб. Получает нервные импульсы от кожи и слизистых желез, преимущественно от зоны иннервации тройничного нерва. Наибольшее значение имеют кожа и слизистые оболочки полости рта и лица, а также твердая мозговая оболочка.
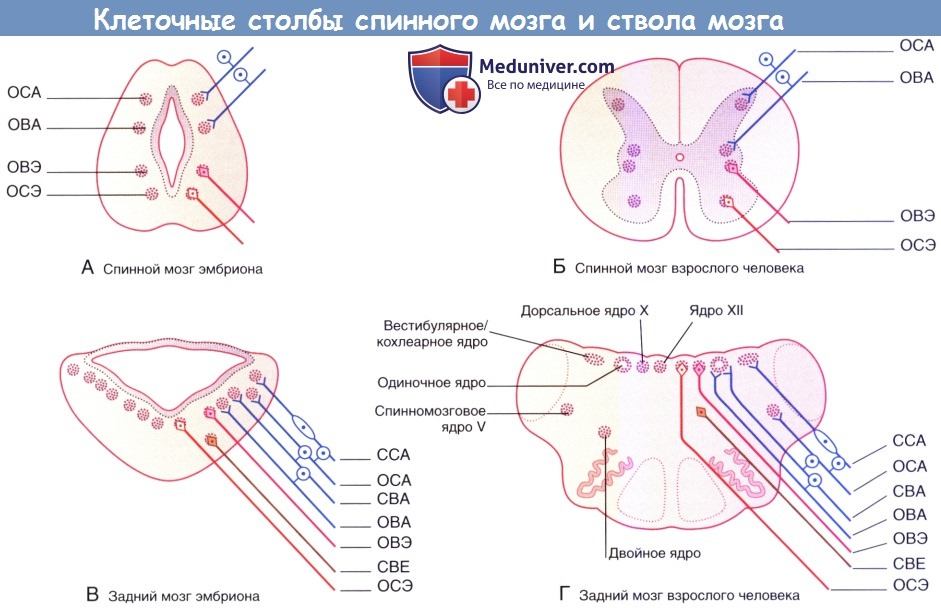
Клеточные столбы спинного мозга и ствола мозга.
(А) Спинной мозг эмбриона. (Б) Спинной мозг взрослого человека.
(В) Задний мозг эмбриона. (Г) Задний мозг взрослого человека.
Афферентные клеточные столбы:
ОСА — общий соматический афферентный; ОВА — общий висцеральный афферентный;
ССА — специальный соматический афферентный; СВА — специальный висцеральный афферентный.
Эфферентные клеточные столбы: ОСЭ — общий соматический эфферентный; ОВЭ — общий висцеральный эфферентный; СВЭ — специальный висцеральный эфферентный.
Дополнительные столбы иннервируют ткани жаберных дуг и внутреннее ухо:
• Специальный висцеральный (бранхиальный) эфферентный (СВЭ) столб. Иннервирует мимическую и жевательную мускулатуру, мышцы неба, глотки и гортани, которые происходят из жаберных дуг. Иннервацию обеспечивают ветви тройничного, лицевого, языкоглоточного, блуждающего и добавочного нервов. Эти поперечно-полосатые мышцы также выполняют висцеральные функции, связанные с употреблением пищи и дыханием (отсюда название — висцеральный столб).
• Специальный висцеральный афферентный (СВА) столб. Получает информацию от вкусовых луковиц, расположенных в энтодермальной выстилке жаберных дуг.
• Специальный соматический афферентный (ССА) столб. Получает нервные импульсы от вестибулярного (равновесие) и улиткового (слух) органов внутреннего уха.
На рисунке отображено расположение различных ядер на дорсальном срезе ствола мозга.
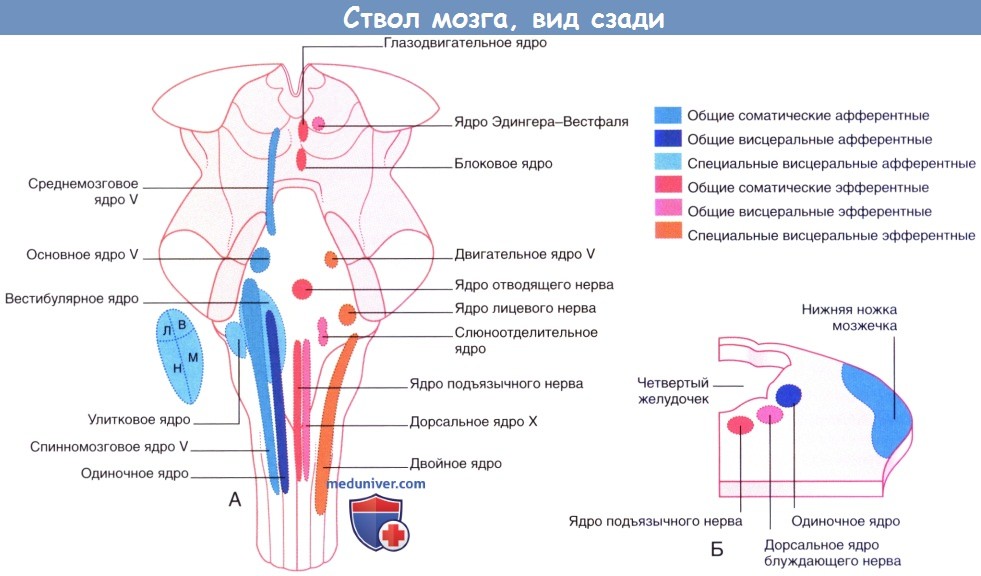
Ствол мозга взрослого человека, вид сзади. Показано положение клеточных столбов черепных нервов.
Л, В, Н, М — латеральное, верхнее, нижнее, медиальное вестибулярные ядра соответственно
(для улучшения информативности рисунка вестибулярные ядра вынесены отдельно).
- Вернуться в оглавление раздела "Неврология."
Редактор: Искандер Милевски. Дата публикации: 16.11.2018
Читайте также:
