
Закон силы длительности в отношении нерва или мышцы проявляется в том что
Все возбудимые клетки (ткани) обладают рядом общих физиологических свойств (законы раздражения), краткая характеристика которых приводится ниже. Универсальным раздражителем для возбудимых клеток является электрический ток.
Простая возбудимая система – это одна возбудимая клетка, которая реагирует на раздражитель как единое целое.
В простых возбудимых системах подпороговые раздражители не вызывают возбуждения, сверхпороговые раздражители вызывают максимальное возбуждение (рис. 1). При подпороговых значениях раздражающего тока возбуждение (ЭП, ЛО) носит местный (не распространяется), градуальный (сила реакции пропорциональная силе действующего стимула) характер. При достижении порога возбуждения возникает ответ максимальной силы (ПД). Амплитуда ответа (амплитуда ПД) не изменяется при дальнейшем увеличении силы раздражителя.
Рис. 1. Зависимость силы реакции простой возбудимой системы (клетки) от силы раздражителя.
ПВ – порог возбуждения
Сложная возбудимая система – система, состоящая из множества возбудимых элементов (мышца включает множество двигательных единиц, нерв – множество аксонов). Отдельные элементы системы имеют неодинаковые пороги возбуждения.
Для сложных возбудимых систем амплитуда ответа пропорциональна силе действующего раздражителя (при значениях силы раздражителя от порога возбуждения самого легковозбудимого элемента до порога возбуждения самого трудновозбудимого элемента) (рис. 2). Амплитуда ответа системы пропорциональна количеству вовлеченных в ответ возбудимых элементов. При возрастании силы раздражителя в реакцию вовлекается все большее число возбудимых элементов.
Рис. 2. Зависимость силы реакции сложной возбудимой системы (нерв, мышца) от силы раздражителя.
ПВ мin – порог возбуждения самого легковозбудимого элемента,
ПВ мах – порог возбуждения самого трудновозбудимого элемента
Эффективность раздражителя зависит не только от силы, но и от времени его действия. Сила раздражителя, вызывающего процесс распространяющегося возбуждения, находится в обратной зависимости от длительности его действия. Графически эта закономерность выражается кривой Вейсса (рис. 3).
Рис. 3. Зависимость пороговой силы раздражителя от времени его действия (закон силы - длительности).
Р – реобаза, ПВ – полезное время, Х – хронаксия
Минимальную силу раздражителя, вызывающую возбуждение, называют реобазой . Наименьшее время, в течение которого должен действовать раздражитель силой в одну реобазу, чтобы вызвать возбуждение, называют полезным временем . Для более точной характеристики возбудимости используют параметр хронаксия. Хронаксия – минимальное время действия раздражителя в 2 реобазы, необходимое для того, чтобы вызвать возбуждение.
Для возникновения возбуждения имеет значение не только сила и время действия тока, но и скорость нарастания силы тока. Для возникновения возбуждения сила раздражающего тока должна нарастать достаточно круто (рис. 4). При медленном нарастании силы тока происходит явление аккомодации – возбудимость клетки снижается. В основе явления аккомодации лежит повышение КУД вследствие постепенной инактивации Na+ -каналов.
Рис. 4. Изменение мембранного потенциала и критического уровня деполяризации при медленном ( А ) и быстром ( Б ) нарастании силы раздражающего тока.
Деполяризация, повышение возбудимости и возникновение возбуждения происходят при действии на клетку выходящего тока . При действии входящего тока происходят противоположные изменения – гиперполяризация и снижение возбудимости, возбуждение не возникает. За направление тока принимают направление от области положительного заряда к области отрицательного заряда.
При внеклеточном раздражении возбуждение возникает в области катода (–). При внутриклеточном раздражении для возникновения возбуждения необходимо, чтобы внутриклеточный электрод имел положительный знак (рис. 5).
Рис. 5. Изменения, наступающие в нервном волокне при внутриклеточном или внеклеточном раздражении.
Стрелкой показано направление электрического тока
Под лабильностью понимают функциональную подвижность, скорость протекания элементарных физиологических процессов в клетке (ткани). Количественной мерой лабильности является максимальная частота циклов возбуждения, которую может воспроизводить клетка. Частота циклов возбуждения не может возрастать беспредельно, так как в каждом цикле возбуждения имеется период рефрактерности. Чем короче рефрактерный период, тем больше лабильность клетки.
Закон силы. Чтобы возникло возбуждение, раздражитель должен быть достаточно сильным — пороговой или сверхпороговой силы. Потенциал действия возникает только при достижении критического уровня деполяризации клеточной мембраны. Критический уровень деполяризации — минимальная деполяризация клеточной мембраны, при которой возникает потенциал действия. Дальнейшее раздражение клетки не изменяет процесс возникновния ПД, так как деполяризация клетки, достигнув критической величины, способствует открытию потенциалозависимых ворот № + -каналов, в результате чего ионы натрия устремляются в клетку, ускоряя деполяризацию независимо от действия раздражителя. Критический уровень деполяризации клеточной мембраны нейрона составляет около —50 мВ. В соответствии с этим законом функционируют поперечнополосатые мышцы. Например, амплитуда сокращений скелетной мышцы постепенно увеличивается с нарастанием силы раздражителя до максимальных значений. Это связано с тем, что скелетные мышцы состоят из множества мышечных волокон, имеющих неодинаковую возбудимость. Поэтому сначала на пороговый раздражитель отвечают те мышечные волокна, которые имеют более высокую возбудимость. С увеличением силы раздражителя в реакцию вовлекается все большее число мышечных волокон, поскольку для менее возбудимых из них эта сила будет пороговой. В результате амплитуда сокращения мышцы увеличивается. После возбуждения всех волокон данной мышцы дальнейшее увеличение силы раздражителя уже не приводит к увеличению амплитуды сокращения.
Закон градиента, или аккомодации. Этот закон гласит, что действие раздражителя зависит не только от абсолютной величины его силы, но и от скорости ее нарастания до порогового значения. Например, действие очень медленно нарастающего раздражения не вызывает возбуждения, так как раздражаемая ткань адаптируется к его влиянию. Этот факт объясняется тем, что при действии медленно возрастающего раздражителя в мембране возбудимой ткани увеличивается уровень деполяризации.
При снижении скорости нарастания силы раздражителя до некоторого минимального значения ответная реакция не возникает при любой силе раздражителя. Это явление получило название аккомодации. Причиной аккомодации является инактивация № + -каналов, возникающая при медленной деполяризации клеточной мембраны, которая длится в течение 1 с и более.
Способность к аккомодации у различных структур неодинакова. Она высокая у нервных волокон и низкая у сердечной мышцы, гладких мышц кишечника и желудка.
Закон длительности силы. Согласно этому закону для проявления реакции на раздражитель необходимо не только применять раздражитель пороговой силы, но и обеспечить его действие в течение определенного времени. Чем больше сила раздражителя, тем менее продолжительно он должен действовать для того, чтобы возникло возбуждение.
Зависимость между силой постоянного электрического тока и длительностью его воздействия для появления ответной реакции имеет гиперболический характер. Поэтому воздействие тока ниже минимальной величины не вызовет возбуждение в ткани, как бы длительно он ни действовал. Кроме того, чем короче импульс тока, тем меньше его раздражающая способность. Причиной этой зависимости является неспособность раздражителя осуществить определенный сдвиг мембранного потенциала вследствие кратковременности действия или недостаточной силы раздражителя.
Минимальная величина тока, способная вызвать возбуждение, называется реобазой. Если возбудимость ткани высока, то пороговая сила раздражителя может быть низкой.
Пороговое время называют также полезным временем. В связи с тем что определить это время достаточно сложно, было введено понятие хронаксии. Хронаксия — минимальное время, в течение которого должен действовать ток в две реобазы, чтобы вызвать возбуждение. Хронаксия измеряется в миллисекундах. Чем меньше полезное время и хронаксия, тем выше возбудимость ткани.
Закон полярного действия постоянного тока. Этот закон проявляется в том, что при действии постоянного тока средней силы на ткань возбуждение возникает только в момент замыкания и в момент размыкания цепи.
Во время замыкания тока возбуждение происходит в участке, расположенном под катодом, а при размыкании — под анодом. Обусловлено это тем, что прохождение постоянного электрического тока через возбудимую ткань вызывает изменение мембранного потенциала покоя ее клеток. В области катода положительный потенциал на наружной поверхности клеточной мембраны уменьшается, и происходит деполяризация мембраны, которая быстро достигает критического уровня и вызывает возбуждение. В области анода положительный потенциал на наружной поверхности мембраны клеток ткани возрастает и развивается гиперполяризация мембраны. При размыкании электрической цепи на аноде мембранный потенциал быстро возвращается к исходному уровню и достигает критической величины, в результате чего открываются ворота № + -каналов и возникает размыкательное возбуждение.
Контрольные вопросы и задания
Законы раздражения отражают определенную зависимость между действием раздражителя и ответной реакцией возбудимой ткани. К законам раздражения относятся, закон силы, закон "все или ничего", закон аккомодации (Дюбуа-Реймона), закон силы-времени (силы-длительности), закон полярного действия постоянного тока, закон физиологического электротона.
1. Закон силы: чем больше сила раздражителя, тем больше величина ответной реакции. В соответствии с этим законом функционируют сложные структуры, например, скелетная мышца. Амплитуда ее сокращений от минимальных (пороговых) величин постепенно увеличивается с увеличением силы раздражителя до субмаксимальных и максимальных значений. Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих различную возбудимость.
Поэтому на пороговые раздражители отвечают только те мышечные волокна, которые имеют самую высокую возбудимость, амплитуд, мышечного сокращения при этом минимальна. С увеличением силы раздражителя в реакцию вовлекается все большее количество мышечных волокон, и амплитуда сокращения мышц все время увеличивается. Когда в реакцию вовлечены все мышечные волокна, составляющие данную мышцу, дальнейшее увеличение силы раздражителя не приводит к увеличению амплитуды сокращения.
3. Закон раздражения - Дюбуа-Реймона (аккомодации) раздражающее действие постоянного тока зависит не только от абсолютной величины силы тока или его плотности, но и от скорости нарастания тока во времени. При действии медленно нарастающего раздражителя возбуждение не возникает, так как происходит приспосабливание возбудимой ткани к действию этого раздражителя, что получило название аккомодации. (Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране возбудимой ткани происходит повышение критического уровня деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения потенциал действия вообще не возникает.
Причина заключается в том, что деполяризация мембраны является пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению натриевой проницаемости, и тем самым обусловливающего возникновение потенциала действия, и медленного, приводящего к инактивации натриевой проницаемости и как следствие этого - окончанию потенциала действия. При быстром нарастании стимула повышение натриевой проницаемости успевает достичь значительной величины прежде, чем наступит инактивация натриевой проницаемости. При медленном нарастании тока на первый план выступают процессы инактивации, приводящие к повышению порога или ликвидации возможности генерировать ПД вообще).
Под градиентом раздражения понимают скорость нарастания силы раздражения до определенной величины. При очень медленном нарастании силы раздражителя порог возбудимости повышается и потенциал действия не возникает, т.е. аккомодация – это увеличение порога возбудимости при действии медленно нарастающей силе раздражителя. Дебуа-Реймон (1818-1896).
Способность к аккомодации различных структур неодинакова. Наиболее высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка.
4. Закон силы-длительности: раздражающее действие постоянного тока зависит не только от его величины, но и от времени, в течение которого он действует. Чем больше ток, тем меньше времени он должен действовать для возникновения возбуждения.
Причиной такой зависимости является мембранная емкость. Очень "короткие" токи просто не успевают разрядить эту емкость до критического уровня деполяризации. Раздражитель, способный вызвать ответную реакцию, называется пороговым. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, названа Лапиком реобазой. Время, в течение которого действует ток, равный реобазе, и вызывает возбуждение, называется полезным временем. Это означает, что дальнейшее увеличение времени не имеет смысла для возникновения потенциала действия (ПД).
В связи с тем, что определение этого времени затруднено, было введено понятие хронаксия - минимальное время, в течение которого ток, равный двум реобазам, должен действовать на ткань, чтобы вызвать ответную реакцию. Определение хронаксии - хронаксиметрия - находит применение в клинике. Электрический ток, приложенный к мышце, проходит через как мышечные, так и нервные волокна и их окончания, находящиеся в этой мышце. Хронаксия нервных и мышечных волокон равна тысячным долям секунд. Если нерв поврежден или произошла гибель соответствующих мотонейронов спинного мозга (это имеет место при полиомиелите и некоторых других заболеваниях), то происходит перерождение нервных волокон и тогда определяется хронаксия уже мышечных волокон, которая имеет большую величину, чем нервных волокон.
Законы раздражения возбудимости тканей
Закон силы
Возникновение распространяющегося возбуждения (ПД) возможно при условии, когда действующий на клетку раздражитель имеет некоторую минимальную (пороговую силу), иначе говоря, когда сила раздражителя соответствует порогу раздражения.
Порог раздражения – это та наименьшая величина раздражителя, которая действуя на клетку какое-то определенное время, способна вызвать максимальное возбуждение.
– наименьшая величина раздражителя, при действии которой потенциал покоя может сместиться до уровня критической деполяризации;
– критическая величина деполяризации клеточной мембраны, при которой активируется перенос ионов натрия внутрь клетки.

Рисунок 5. Возникновение местного потенциала при прохождении электрического тока, через участок нерва. Ток идет от анода к катоду (оба электрода – вне нерва) частично через пленку жидкости на поверхности нерва, а частично через оболочку нерва и в продольном направлении внутри волокна. Кривая внизу показывает вызываемое током изменения мембранного потенциала нервного волокна (по В. Катц)
Зависимость пороговой силы стимула от его длительности
Пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. Эта зависимость, открытая Гоорвегом, Вейсом, Лапиком получила название кривой “сила – длительность” или “сила – время”. Кривая “силы времени” имеет форму, близкую к равносторонней гиперболе и в первом приближении может быть описана эмпирической формулой:
где I – сила тока, Т – длительность его действия, а и b – постоянные, определяемые свойствами ткани.
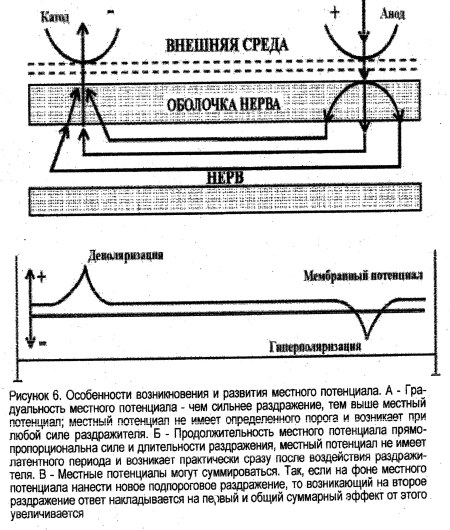
Рисунок 6. Особенности возникновения и развития местного потенциала. А – Градуальность местного потенциала – чем сильнее раздражение, тем выше местный потенциал; местный потенциал не имеет определенного порога и возникает при любой силе раздражителя. Б – Продолжительность местного потенциала прямопропорциональна силе и длительности раздражения, местный потенциал не имеет латентного периода и возникает практически сразу после воздействия раздражителя. В – Местные потенциалы могут суммироваться. Так, если на фоне местного потенциала нанести новое подпороговое раздражение, то возникающий на второе раздражение ответ накладывается на первый и общий суммарный эффект от этого увеличивается
Таким образом, го этой кривой вытекают два следствия:
1. Ток величиной ниже порога не вызывает возбуждения, как бы длительно он не действовал.
2. Какой бы сильный не был раздражитель, но если он действует очень короткое время, то возбуждение не возникает.
Реобаза – минимальная сила тока (или напряжение), способная вызвать возбуждение. Наименьшее время, в течение которого должен действовать стимул в одну реобазу, чтобы вызвать возбуждение – полезное время. Дальнейшее его увеличение не имеет значения для возникновения возбуждения.
Порог (реобаза) – величины непостоянные, зависят от функционального состояния клеток в покое, поэтому Лапик предложил определять более точный показатель – хронаксию.
Хронаксия – наименьшее время, в течение которого ток в две реобазы должен действовать на ткань, чтобы вызывать возбуждение.
Метод определения хронаксии – хронаксиметрия используется в клинике для диагностики повреждения нервных стволов и мышц.
Зависимость порога от крутизны нарастания раздражителя (аккомодация)
Порог раздражения имеет наименьшую величину при толчках электрического тока прямоугольной формы, когда сила нарастает очень быстро.
При уменьшении крутизны нарастания стимула ускоряются процессы инактивации натриевой проницаемости, приводящие к повышению порога и снижению амплитуды потенциалов действия. Чем круче должен нарастать ток, чтобы вызвать возбуждение, тем выше скорость аккомодации. Очень мала скорость аккомодации тех образований, которые склонны к автоматической деятельности (миокард, гладкие мышцы).
“Все” — на пороговые и сверхпороговые стимулы ответ максимальный и развивается потенциал действия; “ничего” – надо – пороговый стимул потенциал действия не развивается. Закон “все или ничего” установлен Боудичем в 1871 г. на мышце сердца: при подпороговой силе раздражения мышца сердца не сокращается, а при пороговой силе раздражения – сокращение максимально. При дальнейшем увеличении силы раздражения амплитуда сокращений не увеличивается.
Со временем была установлена и относительность этого закона. Оказалось, что “все” зависит от функционального состояния ткани (охлаждение, исходное растяжение мышцы и т. д.). С появлением микроэлектродной техники было установлено еще одно несоответствие: подпороговое раздражение вызывает местное, не распространяющееся возбуждение, следовательно, нельзя говорить, что допороговое раздражение не дает ничего. Процесс развития возбуждения подчиняется этому закону с уровня критической деполяризации, когда запускается лавинообразное поступление ионов калия в клетку.
Изменение возбудимости при возбуждении
Мера возбудимости – это порог раздражения. При местном, локальном, возбуждений возбудимость увеличивается. Потенциал действия сопровождается многофазными изменениями возбудимости.
1. Период повышенной возбудимости соответствует локальному ответу, когда мембранный потенциал достигает УКД, возбудимость повышена.
2. Период абсолютной рефрактерности соответствует фазе деполяризации потенциала действия, пику и началу фазы реполяризации, возбудимость снижена вплоть до полного отсутствия во время пика.
3. Период относительной рефрактерности соответствует оставшейся части фазы реполяризации, возбудимость постепенно восстанавливается к исходному уровню.
4. Супернормальный период соответствует фазе следовой деполяризации потенциала действия (отрицательный следовый потенциал), возбудимость повышена.
5. Субнормальный период соответствует фазе следовой гиперполяризации потенциала действия (положительный следовый потенциал), возбудимость снижена.

Рисунок 7. Изменения возбудимости нервного волокна в различные фазы потенциала действия и следовых изменений потенциала действия (по Б.И. Ходорову). Для наглядности длительность первых двух фаз на каждой кривой несколько увеличена. Пунктирной линией на рисунке А обозначает потенциал покоя, а на рисунке Б исходный уровень возбудимости
Закон лабильности или функциональной подвижности
Лабильность — скорость протекания физиологических процессов в возбудимой ткани. Например, можно творить о максимальной частоте раздражения, которую возбудимая ткань способна воспроизводить без трансформации ритма.
Мерой лабильности могут служить:
– длительность отдельного потенциала
– величина абсолютной рефрактерной фазы
– скорость восходящей и нисходящей фаз ПД.
Уровень лабильности характеризует скорость возникновения и компенсации возбуждения в любых клетках и уровень их функционального состояния. Можно измерять лабильность мембран, клеток, органов, причем, в системе из нескольких элементов (тканей, органов, образований) лабильность определяется по участку с наименьшей лабильностью:
Полярный закон раздражения (закон Пфлюгера)
Закон изменения мембранного потенциала при действии на возбудимые ткани постоянного электрического тока открыл Пфлюгер в 1859 г.
1. Постоянный ток проявляет свое раздражающее действие только в момент замыкания и размыкания цепи.
2. При замыкании цепи постоянного тока возбуждение возникает под катодом; при размыкании по анодом.
Изменение возбудимости под катодом.
При замыкании цепи постоянного тока под катодом (действуют допороговым, но продолжительным раздражителем) на мембране возникает стойкая длительная деполяризация, которая не связана с изменением ионной проницаемости мембраны, а обусловлена перераспределением ионов снаружи (они привносятся на электроде) и внутри – катион перемещается к катоду.
Вместе со смещением мембранного потенциала уровень критической деполяризации (УКД) смещается к нулю. При размыкании цепи постоянного тока под катодом мембранный потенциал быстро возвращается к исходному уровню, а УКД медленно, следовательно, порог увеличивается, возбудимость снижается и отмечается католическая депрессия Вериго. Таким образом, возбуждение возникает только при замыкании цепи постоянного тока под катодом.
Изменение возбудимости под анодом.
При замыкании цепи постоянного тока под анодом (действуют допороговым, но продолжительным раздражителем) на мембране развивается гиперполяризация за счет перераспределения ионов по обе стороны мембраны (без изменения ионной проницаемости мембраны) и возникающее за ней смещение уровня критической деполяризации в сторону мембранного потенциала. Следовательно, порог уменьшается, возбудимость повышается – анодическая экзальтация.
При размыкании цепи мембранный потенциал быстро восстанавливается к исходному уровню и достигает сниженного уровня критической деполяризации, генерируется потенциал действия. Таким образом, возбуждение возникает только при размыкании цепи постоянного тока под анодом. Сдвиги мембранного потенциала вблизи полюсов постоянного тока получили название электротонических. Сдвиги мембранного потенциала не связанные с изменением ионной проницаемости мембраны клетки называют пассивными.
Проведение возбуждения.
Потенциал действия – это волна возбуждения, распространяющаяся по мембранам нервных и мышечных клеток.
ПД обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам. Синоним ПД – нервный импульс или спайк. Сложная информация о действующих на организм раздражениях кодируется в виде отдельных групп потенциалов действия – рядов.
Согласно закону “все или ничего” амплитуда и длительность отдельных потенциалов действия постоянны, а частота и количество в ряду зависит от интенсивности раздражения. Такой способ кодирования информации и ее передачи является наиболее психоустойчивым.
В живых организмах информация может передаваться и гуморальным путем.
Преимущества ПД:
1. Информация более целенаправлена;
2. Передается быстро;
3. Адресат точно известен;
4. Информация может быть точнее закодирована.
ПД распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками. Из-за перезарядки мембраны во время генерации потенциала действия последний обладает способностью к самораспространению. Возникнув на одном участке, является стимулом для соседних. Наступающая после возбуждения в данном участке мембраны рефрактерностъ, обусловливает поступательное движение ПД.
Конкретные особенности распространения возбуждения связаны со строением мембраны клетки, нервных волокон. По мембранам мышечных клеток и в безмякотных нервных волокнах возбуждение распространяется непрерывно вдоль всей мембраны.
В волокнах, покрытых миелиновой оболочкой, потенциал действия может распространяться только скачкообразно (сальтаторно), перепрыгивая через участки волокна, покрытые шванновскими клетками, с одного перехвата Ранвье на другой.
Перехваты Ранвье представляют собой своеобразные станции ретрансляции, постоянно усиливающие сигнал, не давая ему угаснуть.
Причины сальтаторного проведения:
1. В перехватах Ранвье, свободных от миелина, сопротивление электрическому току минимально;
2. Порог раздражения в перехватах Ранвье минимальный;
3. Амплитуда ПД в каждом перехвате в 5 – 6 раз превышает порог в соседнем перехвате;
4. Велика плотность натриевых каналов на мембране перехвата.
Следовательно, возбуждение, возникающее в одном перехвате Ранвье, вызывает смещение электронов во внешней среде данного волокна и этого смещения достаточно, чтобы вызвать возбуждение в соседнем участке. Таким образом, скорость проведения возбуждения по нервному волокну зависит от диаметра волокон и наличия перехватов Ранвье.
Различают декрементное и бездекрементное распространение волны возбуждения.
ДЕКРЕМЕНТНОЕ проведение:
1. Наблюдается в безмиелиновых волокнах;
2. Скорость проведения невелика;
3. По мере удаления от места возникновения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания;
4. Свойственно волокнам, которые иннервируют внутренние органы, обладающие низкой функциональной активностью.
БЕЗДЕКРЕМЕНТНОЕ проведение:
1. ПД проходит весь путь от места раздражения до места реализации без затухания.
2. Характерно для миелиновых и тех безмиелиновых волокон, которые передают сигналы к органам, обладающими высокой реактивностью (сердце).
Распространение одиночного потенциала действия само по себе не требует энергетических затрат. Однако, восстановление исходного состояния мембраны и поддержание ее готовности к проведению нового импульса связано с затратой энергии.
Законы проведения возбуждения в нервах
Закон анатомической и физиологической непрерывности волокна.
Любая травма волокна нарушает проводимость. При действии новокаина (дикаина, кокаина) блокируются натриевые и калиевые каналы мембраны. Возникновение возбуждения и его проведение в этом случае становится невозможным.
Закон двустороннего проведения возбуждения
В целом организме по рефлекторной дуге возбуждение всегда распространяется в одном направлении: от рецептора к эффектору.
Причины:
1. Возбуждение всегда возникает при раздражении специфических рецепторов;
2. Рефрактерность во время возбуждения обусловливает поступательное движение;
3. В рефлекторной дуге возбуждение с одной нервной клетки на другую передастся в синапсах с помощью медиатора, который может выделяться только в одном направлении.
Закон изолированного проведения возбуждения в нервных стволах.
Передача возбуждения на большие расстояния невозможна из-за значительной потери тока во внеклеточной среде.
Физиология нейронов, глиальных клеток, рецепторов и синапсов
Классическая рефлекторная дуга состоит из:
– рецептора;
– афферентного пути (афферентного нейрона, который располагается в спинальном ганглии);
– нервного центра, где возбуждение с афферентного нейрона переходит на вставочную нервную клетку.
Затем возбуждение переходит на эффекторный орган (эффектор), в роли которого может выступать мышца. Многие нервные волокна покрыты глиальными клетками (миелиновая оболочка). Между этими Шванновскими клетками есть промежутки – перехваты Ранвье. Возбуждение с одного нейрона на другой и с мотонейрона на мышцу передается в синапсах с помощью медиатора.
Нервная клетка – структурная и функциональная единица ДНС, которая окружена клетками нейроглии.
Нейроглия (глиоциты) – совокупность всех клеточных элементов нервной ткани кроме нейронов.
В мозге взрослого человека 1150 – 200 млрд. глиальных клеток, что в 10 раз больше нервных.
Нейроглия:
1. макроглия:
– астроциты;
– олигодендроциты;
– эиендимоциты.
2. микроглия: глиальные макрофаги.
Астроциты составляют 45 – 60% серого вещества мозга. Покрывают 85% поверхности капилляров мозга (сосудистые ножки астроцитов), крупные отростки астроцитов контактируют с телами нейронов. Основная функция – трофическая.
Олигодендроциты образуют миелин в нервной системе к поддерживают его целостность.
Эпендимоциты – клетки, выстилающие стенки спинномозгового канала и всех желудочков головного мозга. Это граница между спинномозговой жидкостью (ликвор) и тканью мозга.
Функции нейроглии:
1. Опорная – вместе с сосудами и мозговыми оболочками образуют строму ткани мозга.
2. Трофическая – обеспечивают метаболизм нервных клеток (связь с кровеносными сосудами). В глиоцитах сосредоточен весь гликоген ЦНС.
3. Участие в интегративной деятельности мозга:
– формирование следов воздействия (память), а значит и условного рефлекса;
– без глиоцитов (блокада антиглиальным гамма-глобулином) меняется электрическая активность нейронов.
Особенности глиальных клеток:
1. Более чувствительны к ионным изменениям среды;
2. Высокая активность калий – натриевой АТФ-азы;
3. Высокая проницаемость для ионов калия;
4. Мембранный потенциал равен 90 мВ, у нейронов 60 – 80 мВ;
5. На раздражение отвечает только медленной деполяризацией не более 10 мВ;
6. Потенциал действия в глиальных клетках не генерируется.
Читайте также:
