
Зона невозбудимости аксона после проведения нервного импульса это
Возбуждение в виде потенциала действия покидает тело нейрона по его отростку, который называется аксоном. Аксоны отдельных нейронов обычно объединяются в пучки — нервы, а сами аксоны в этих пучках называются нервными волокнами. Природа позаботилась, чтобы волокна максимально хорошо справлялись с функцией проведения возбуждения в виде потенциалов действия. Для этой цели отдельные нервные волокна (аксоны отдельных нейронов) имеют специальные чехлы, выполненные из хорошего электрического изолятора (см. рис. 2.3). Чехол прерывается примерно через каждые 0,5-1,5 мм; это связано с тем, что отдельные участки чехла образуются в результате того, что специальные клетки в очень ранний период развития организма (в основном еще до рождения) обволакивают небольшие участки аксона. На рис. 2.9 показано, как это происходит. В периферических нервах миелин
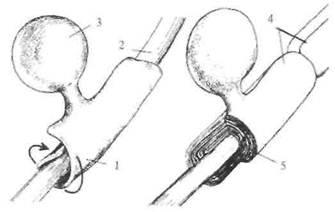
Рис. 2.9. Образование миелинового чехла на аксоне.
1 — наматывание слоев миелина; 2 — аксон: 3 — олигодендроцит; 4 — перехват Ранвье; 5 — слой миелиновой оболочки.
образуется клетками, которые получили название шванновских, а в головном мозге это происходит за счет клеток олигодендроглии. Этот процесс называется миелинизацией, так как в результате образуется чехол из вещества миелина, примерно на 2 /3 состоящего из жира и являющегося хорошим электрическим изолятором. Исследователи придают очень большое значение процессу миелинизации в развитии мозга.
Известно, что у новорожденного ребенка миелинизировано примерно 2 /3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций. В то же время известны заболевания нервной системы человека, которые связаны с демиелинизацией нервных волокон, что сопровождается тяжелыми страданиями. К самым известным относится рассеянный склероз. Это заболевание развивается незаметно и очень медленно, последствием является паралич движения.
Почему же так важна миелинизация нервных волокон? Оказывается, миелинизированные волокна в сотни раз быстрее проводят
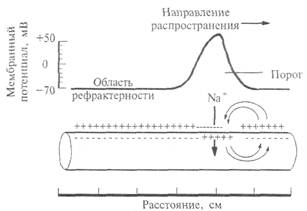
Рис. 2.10. Распространение возбуждения по немиелинизированному нервному волокну: после прохождения потенциала действия в нервном волокне возникает зона невозбудимости, или рефрактерности.
возбуждение, чем немиелинизированные, т. е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно. Поэтому не миелинизируются в нашем организме только самые тонкие волокна (менее 1 мкм в диаметре), которые проводят возбуждение к медленно работающим органам — кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.
Как происходит распространение возбуждения по нервному волокну? Вначале разберем случай немиелинизированного нервного волокна. На рис. 2.10 показана схема нервного волокна. Возбужденный участок аксона характеризуется тем, что мембрана, обращенная к аксоплазме, заряжается положительно относительно экстраклеточной среды. Невозбужденные (покоящиеся) участки мембраны волокна отрицательны внутри. Между возбужденным и невозбужденным участками мембраны возникает разность потенциалов и начинает протекать ток. На рисунке это отражено линиями тока, пересекающими мембрану со стороны аксоплазмы, — выходящий ток, который деполяризует соседний невозбуждениый участок волокна. Возбуждение движется по волокну только в одном направлении (показано стрелкой) и не может пойти в другую сторону, так как после возбуждения участка волокна в нем наступает рефрактерность — зона не возбудимости. Нам уже известно,
Рис. 2.11. Распространение возбуждения по миелинизированному нервному волокну: потенциалы действия возникают только в перехватах Ранвье.
что деполяризация приводит к открыванию потенциалзависимых натриевых каналов и в соседнем участке мембраны развивается потенциал действия. Затем натриевый канал инактивируется и закрывается, что и приводит к зоне невозбудимости волокна. Эта последовательность событий повторяется для каждого соседнего участка волокна. На каждое такое возбуждение тратится определенное время. Специальные исследования показали, что скорость проведения возбуждения немиелинизированных волокон пропорциональна их диаметру: чем больше диаметр, тем выше скорость движения импульсов. Например, немиелинизированные волокна, проводящие возбуждение со скоростью 100—120 м/с, должны иметь диаметр около 1000 мкм (1 мм).
У млекопитающих животных природа сохранила немиелинизированными только те волокна, которые проводят возбуждение о боли, температуре, управляют медленно работающими внутренними органами — мочевым пузырем, кишечником и пр. Практически все нервные волокна в центральной нервной системе человека имеют миелиновые чехлы. На рис. 2.11 показано, что если вдоль волокна, покрытого миелином, регистрировать прохождение возбуждения, то потенциал действия возникает только в перехватах Ранвье. Оказывается, миелин, являясь хорошим электрическим изолятором, не пропускает выхода линий тока от предшествующего возбужденного участка. Выход тока в этом случае возможен только через те участки мембраны, которые находятся на стыке между двумя участками миелина. Напомним, что каждый участок образован только одной клеткой, поэтому это стыки между двумя клетками, образующими соседние участки миелиновой оболочки. Мембрана аксона между двумя соседними миелиновыми чехлами оказывается не покрытой миелином (так называемый перехват Ранвье). Благодаря такому устройству мембрана волокна возбуждается только в местах перехватов Ранвье. Вследствие этого потенциал действия (возбуждение) как бы перескакивает через участки изолированной мембраны. Другими словами, возбуждение движется скачками от перехвата к перехвату. Это похоже на те волшебные сапоги-скороходы, которые надевал кот в известной сказке, мгновенно переносясь из одного места в другое.
СИНАПС
Как передается возбуждение — от одного нейрона другому или от нейрона, например, на мышечное волокно? Этой проблемой интересуются не только профессиональные нейробиологи, но и врачи, особенно фармакологи. Знание биологических механизмов необходимо для лечения некоторых заболеваний, а также для создания новых лекарств и препаратов. Дело в том, что одними из основных мест воздействия этих веществ на организм человека являются места передачи возбуждения с одного нейрона на другой (или на другую клетку, например клетку сердечной мышцы, стенки сосудов и пр.). Отросток нейрона аксон направляется к другому нейрону и образует на нем контакт, который называют синапсом (в переводе с греческого — контакт; см. рис. 2.3). Именно синапс хранит многие тайны мозга. Нарушение этого контакта, например, веществами, блокирующими его работу, приводит к тяжелейшим последствиям для человека. Это место приложения действия наркотиков. Примеры будут приведены ниже, а сейчас рассмотрим, как устроен и как работает синапс.
Трудности этого исследования определяются тем, что сам синапс очень маленький (его диаметр не более 1 мкм). Один нейрон получает такие контакты, как правило, от нескольких тысяч (3— 10 тыс.) других нейронов. Каждый синапс надежно закрыт специальными
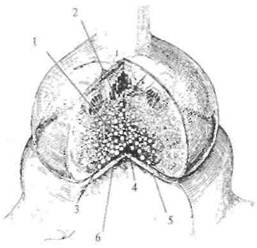
Рис. 2.12. Синапс (схема).
1 — везикулы; 2 — митохондрии; 3 — пресинаптическая мембрана; 4 — синаптическая щель; 5 — ионный канал; 6 — постсинаптическая мембрана.
клетками глии, поэтому исследовать его очень непросто. На рис. 2.12 показана схема синапса, как это представляет себе современная наука. Несмотря на свою миниатюрность, он устроен весьма сложно. Одним из его основных компонентов являются пузырьки, которые находятся внутри синапса. Эти пузырьки содержат биологически очень активное вещество, которое называется нейротрансмиттером, или медиатором (передатчиком).
Вспомним, что нервный импульс (возбуждение) с огромной скоростью продвигается по волокну и подходит к синапсу. Этот потенциал действия вызывает деполяризацию мембраны синапса (рис. 2.13), однако это не приводит к генерации нового возбуждения (потенциала действия), а вызывает открывание специальных ионных каналов, с которыми мы еще не знакомы. Эти каналы пропускают ионы кальция внутрь синапса. Ионы кальция играют очень большую роль в деятельности организма. Специальная железа внутренней секреции — паращитовидная (она находится поверх щитовидной железы) — регулирует содержание кальция в организме. Многие заболевания связаны с нарушением обмена кальция в организме. Например, его недостаток приводит к рахиту у маленьких детей.
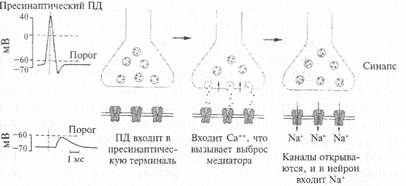
Рис. 2.13. Последовательность событий при возбуждении синапса.
На мембране одного нейрона могут одновременно находиться два вида синапсов: тормозные и возбудительные. Все определяется устройством ионного канала мембраны. Мембрана возбудительных синапсов пропускает как ионы натрия, так и ионы калия. В этом случае мембрана нейрона деполяризуется. Мембрана тормозных синапсов пропускает только ионы хлора и гиперполяризуется. Очевидно, что если нейрон заторможен, потенциал мембраны увеличивается (гиперполяризация). Таким образом, нейрон благодаря воздействию через соответствующие синапсы может возбудиться или прекратить возбуждение, затормозиться. Все эти события происходят на соме и многочисленных отростках дендрита нейрона, на последних находится до нескольких тысяч тормозных и возбудительных синапсов.
В качестве примера разберем, как действует в синапсе медиатор, который называется ацетилхолином. Этот медиатор широко распространен в головном мозге и в периферических окончаниях нервных волокон. Например, двигательные импульсы, которые по соответствующим нервам приводят к сокращению мышц нашего тела, оперируют ацетилхолином. Ацетилхолин был открыт в 30-х годах австрийским ученым О. Леви. Эксперимент был очень прост: изолировали сердце лягушки с подходящим к нему блуждающим нервом. Было известно, что электрическая стимуляция блуждающего нерва приводит к замедлению сокращений сердца вплоть до полной его остановки. О. Леви простимулировал блуждающий нерв, получил эффект остановки сердца и взял из сердца немного крови.
Оказалось, что если эту кровь добавить в желудочек работающего сердца, то оно замедляет свои сокращения. Был сделан вывод: при стимуляции блуждающего нерва выделяется вещество, останавливающее сердце. Это и был ацетилхолин. Позже был открыт фермент, который расщеплял ацетилхолин на холин (жир) и уксусную кислоту, в результате чего прекращалось действие медиатора. Этим исследованием впервые была установлена точная химическая формула медиатора и последовательность событий в типичном химическом синапсе. Эта последовательность событий сводится к следующему.
Потенциал действия, пришедший по пресинаптическому волокну к синапсу, вызывает деполяризацию, которая включает кальциевый насос, и ионы кальция поступают в синапс; ионы кальция связываются белками мембраны синаптических пузырьков, что приводит к активному опорожнению (экзоцитозу) пузырьков в синаптическую щель. Молекулы медиатора связываются (узнающим центром) соответствующими рецепторами постсинаптической мембраны, при этом открывается ионный канал. Через мембрану начинает протекать ионный ток, что приводит к возникновению на ней постсинаптического потенциала. В зависимости от характера открытых ионных каналов возникает возбудительный (открываются каналы для ионов натрия и калия) или тормозной (открываются каналы для ионов хлора) постсинаптический потенциал.
Ацетилхолин весьма широко распространен в живой природе. Например, он находится в стрекательных капсулах крапивы, в стрекательных клетках кишечнополостных животных (например, пресноводной гидры, медузы) и пр. В нашем организме ацетилхолин выбрасывается в окончаниях двигательных нервов, управляющих мышцами, из окончаний блуждающего нерва, который управляет деятельностью сердца и других внутренних органов. Человек давно знаком с антагонистом ацетилхолина — это яд кураре, которым пользовались индейцы Южной Америки при охоте на животных. Оказалось, что кураре, попадая в кровь, вызывает обездвиживание животного, и оно погибает фактически от удушья, но кураре не останавливает сердце. Исследования показали, что в организме существуют два типа рецепторов к ацетилхолину: один успешно связывает никотиновую кислоту, а другой — мускарин (вещество, которое выделено из гриба рода Muscaris). На мышцах нашего тела находятся рецепторы никотинового типа к ацетилхолину, тогда как на сердечной мышце и нейронах головного мозга — рецепторы к ацетилхолину мускаринового типа.
К биогенным аминам относят также адреналин и норадреналин, которые содержатся в синапсах нейронов автономной вегетативной нервной системы. Во время стресса под влиянием специального гормона — адренокортикотропного (подробнее см. ниже) — из клеток коры надпочечников в кровь также выбрасываются адреналин и норадреналин.
Возбуждение в виде потенциала действия покидает тело нейрона по его отростку, который называется аксоном. Аксоны отдельных нейронов обычно объединяются в пучки – нервы, а сами аксоны в этих пучках называются нервными волокнами. Природа позаботилась, чтобы волокна максимально хорошо справлялись с функцией проведения возбуждения в виде потенциалов действия. Для этой цели отдельные нервные волокна (аксоны отдельных нейронов) имеют специальные чехлы, выполненные из хорошего электрического изолятора (см. рис. 2.3). Чехол прерывается примерно через каждые 0,5–1,5 мм; это связано с тем, что отдельные участки чехла образуются в результате того, что специальные клетки в очень ранний период развития организма (в основном еще до рождения) обволакивают небольшие участки аксона. На рис. 2.9 показано, как это происходит. В периферических нервах миелин образуется клетками, которые получили название шванновских, а в головном мозге это происходит за счет клеток олигодендроглии.

Этот процесс называется миелинизацией, так как в результате образуется чехол из вещества миелина, примерно на 2/3 состоящего из жира и являющегося хорошим электрическим изолятором. Исследователи придают очень большое значение процессу миелинизации в развитии мозга.
Известно, что у новорожденного ребенка миелинизировано примерно 2/3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций. В то же время известны заболевания нервной системы человека, которые связаны с демиелинизацией нервных волокон, что сопровождается тяжелыми страданиями. К самым известным относится рассеянный склероз. Это заболевание развивается незаметно и очень медленно, последствием является паралич движения.
Почему же так важна миелинизация нервных волокон? Оказывается, миелинизированные волокна в сотни раз быстрее проводят возбуждение, чем немиелинизированные, т.е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно. Поэтому не миелинизируются в нашем организме только самые тонкие волокна (менее 1 мкм в диаметре), которые проводят возбуждение к медленно работающим органам кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.

Как происходит распространение возбуждения по нервному волокну? Вначале разберем случай немиелинизированного нервного волокна. На рис. 2.10 показана схема нервного волокна. Возбужденный участок аксона характеризуется тем, что мембрана, обращенная к аксоплазме, заряжается положительно относительно экстраклеточной среды. Невозбужденные (покоящиеся) участки мембраны волокна отрицательны внутри. Между возбужденным и невозбужденным участками мембраны возникает разность потенциалов и начинает протекать ток. На рисунке это отражено линиями тока, пересекающими мембрану со стороны аксоплазмы, – выходящий ток, который деполяризует соседний невозбужденный участок волокна. Возбуждение движется по волокну только в одном направлении (показано стрелкой) и не может пойти в другую сторону, так как после возбуждения участка волокна в нем наступает рефрактерность – зона невозбудимости. Нам уже известно, что деполяризация приводит к открыванию потенциалзависимых натриевых каналов и в соседнем участке мембраны развивается потенциал действия. Затем натриевый канал инактивируется и закрывается, что и приводит к зоне невозбудимости волокна. Эта последовательность событий повторяется для каждого соседнего участка волокна. На каждое такое возбуждение тратится определенное время. Специальные исследования показали, что скорость проведения возбуждения немиелинизированных волокон пропорциональна их диаметру: чем больше диаметр, тем выше скорость движения импульсов. Например, немиелинизированные волокна, проводящие возбуждение со скоростью 100–120 м/с, должны иметь диаметр около 1000 мкм (1 мм).
У млекопитающих животных природа сохранила немиелинизированными только те возбуждение о боли, температуре, управляют медленно работающими внутренними органами мочевым волокна, которые проводят органами – мочевым пузырем, кишечником и пр. Практически все нервные волокна в центральной нервной системе человека имеют миелиновые чехлы. На рис. 2.11 показано, что если вдоль волокна, покрытого миелином, регистрировать прохождение возбуждения, то потенциал действия возникает только в перехватах Ранвье. Оказывается, миелин, являясь хорошим электрическим изолятором, не пропускает выхода линий тока от предшествующего возбужденного участка. Выход тока в этом случае возможен только через те участки мембраны, которые находятся на стыке между двумя участками миелина. Напомним, что каждый участок образован только одной клеткой, поэтому это стыки между двумя клетками, образующими соседние участки миелиновой оболочки. Мембрана аксона между двумя соседними миелиновыми чехлами оказывается не покрытой миелином (так называемый перехват Ранвье). Благодаря такому устройству мембрана волокна возбуждается только в местах перехватов Ранвье. Вследствие этого потенциал действия (возбуждение) как бы перескакивает через участки изолированной мембраны. Другими словами, возбуждение движется скачками от перехвата к перехвату. Это похоже на те волшебные сапоги-скороходы, которые надевал кот в известной сказке, мгновенно переносясь из одного места в другое.

Данный текст является ознакомительным фрагментом.
У позвоночных животных в процессе эволюции сформировались аксоны, покрытые миелиновой оболочкой, что обеспечило большие скорости проведения нервных импульсов при значительно меньших диаметрах таких аксонов (см. табл. 11.3).
Строение миелинизированного аксона схематично представлено на рис. 11.6, а. Участки аксона длиной в 2-3 мм

На участках аксона, покрытых миелином, мембрана полностью изолирована и не имеет контакта с межклеточной жидкостью, содержащей ионы Na + , вследствие чего потенциал действия здесь сформироваться не может, даже если потенциал аксоплазмы превышает критический потенциал возбуждения Екр. Потенциал действия может сформироваться только в перехватах Ранвье, где мембрана имеет необходимый контакт с межклеточной жидкостью.
Допустим, что один из перехватов Ранвье возбужден, т.е. деполяризован до потенциала фтах. Тогда под действием разности потенциалов (фтах - ф0) между возбужденным и невозбужденным участками в аксоплазме и на наружной стороне мембраны возникают локальные токи, благодаря которым мембранный потенциал распространяется вдоль аксона как по электрическому кабелю и с большой скоростью. Но по мере удаления от возбужденного участка мембранный потенциал аксона экспоненциально уменьшается (рис. 11.6, б), как и любой местный потенциал [см. формулу (11.7)].
Когда этот затухающий мембранный потенциал достигнет ближайшего перехвата Ранвье и повысит там фм до Екр, в мембране этого перехвата открываются Ма + -каналы и генерируется полноценный потенциал действия, что приводит к увеличению мембранного потенциала до максимального значения фтах. Далее весь процесс повторяется, при этом потенциал действия последовательно генерируется лишь в перехватах Ранвье, а на миелинизированных участках аксона происходит его распространение с затуханием, но с большой скоростью, благодаря чему и достигается высокая скорость передачи возбуждения по такому нервному волокну.
Проведение нервного импульса в миелинизированном аксоне называют сапътаторным (скачкообразным), так как потенциал действия генерируется только в перехватах Ранвье и в процессе распространения как бы перескакивает с одного перехвата Ранвье на последующий. Как и в случае безмиели- новых нервных волокон, скорость распространения потенциала действия в миелинизированном аксоне возрастает с увеличением его диаметра, что хорошо видно из табл. 11.3.
Одной из причин высокой скорости проведения потенциала действия в миелинизированных аксонах является высокая плотность Na + -каналов в перехватах Ранвье, где она примерно в 100 раз выше, чем в безмиелиновых волокнах.
Высокая скорость проведения и малый диаметр миелинизированных аксонов обеспечили позвоночным животным возможность создания большого количества параллельных быстропроводящих нервных путей, что способствовало высокой надежности передачи информации по нервной системе.
Если входящий в клетку натриевый ток уменьшить, например, путем снижения концентрации Na + в растворе, окружающем аксон, либо путем инактивации (ухудшения проводимости) Na + -каналов (введением ядов либо воздействием местноанестезирующего препарата), то скорость проведения потенциала действия снижается и в конечном итоге проведение может быть блокировано.
Если с помощью определенных ядов (например, тетродо- токсина) заблокировать натриевые каналы в одном из перехватов Ранвье, то потенциал действия сформируется не на нем, а на следующем перехвате, так как амплитуда затухающего местного потенциала, распространяющегося за счет локальных токов (пунктирная кривая на рис. 11.6, б), еще сможет достичь на нем порогового потенциала возбуждения. Но если заблокировать подряд два или более перехватов Ранвье, то потенциал действия на последующих перехватах Ранвье сформироваться не сможет и проведение нервного импульса будет блокировано.
может возникнуть в следующих ситуациях;
В клинической практике такой механизм, т.е. понижение проницаемости мембран для Na + и сохранение ее или даже повышение для К + , лежит в основе часто встречающейся патологии — нарушения проводимости в сердечных нервах — в частности, в возникновении периодов Венкенбаха-Самойлова;
— нарушение процесса реполяризации мембраны в результате закрытия Να'-каналов, приводящее к стойкой деполяризации мембраны. К веществам, вызывающим такой эффект, относятся инсектициды (например, ДДТ), ве- ратридин, аконитин, батрахотоксин и др.;
— дефицит энергии ведет к нарушению работы Να*/К*-насоса, что обусловливает неспособность мембраны генерировать потенциал действия и проводить возбуждение. Такой эффект оказывают разобщители окислительного фосфорилирования (например, динитрофенол) и другие метаболические яды, а также ишемия и длительное охлаждение участка нерва. Ингибируют Na + /K + -Hacoc и, как следствие, нарушают проводимость сердечные гликозиды (например, уабаин, строфантин) при применении в больших дозах;
— перехваты Ранвье особенно чувствительны к патогенным блокирующим агентам: кокаин, уретан, ультрафиолетовые лучи, охлаждение и др.;
— повреждение (распад) миелина при дегенерации мякотного нервного волокна или при демиелинизации его при болезнях (рассеянный склероз, вакцинальный процесс, аллергический полиомиелит и др.) приводят к замедлению проведения возбуждения и к дальнейшему нарушению этого процесса вплоть до полного прекращения проводимости. Интересно, что существует минимальное расстояние, на котором повреждение нерва (кроме перерезки) существенно не отражается на функции нейрона. Это расстояние равно расстоянию между двумя соседними перехватами Ранвье — т.е. 1 мм, поскольку, если невозбудимый участок меньше, то потенциал действия, хоть и ослабленный в результате пассивного кабельного проведения, все еще сохранит на другом конце этого участка достаточную силу, чтобы вновь возбудить мембрану и продолжить свой путь;
— развитие парабиоза. Учение о парабиозе было создано Николаем Евгеньевичем Введенским. При проведении импульсов большое значение имеет частота подаваемых на нерв импульсов. Так после возникновения нервного импульса в нерве отмечается абсолютный рефрактерный период, во время которого никакое повторное раздражение не может вызвать второго импульса. Этот период имеет такую же длительность, как и основная часть потенциала действия. Затем наступает относительно рефрактерный период, во время которого можно вызвать второй импульс, но с помощью более сильного раздражителя.
Методика классических опытов с парабиозом несложная. Опыты поставлены на нервно-мышечном препарате (см. рис.). На участок нервного волокна оказывали действие различные патогенные агенты. Выше этого участка фиксировались электроды для раздражения непоражаемого участка нерва. Между этими элетродами и мышцей помещались электроды, отводящие биотоки к регистрирующему аппарату. Благодаря этой методике удалось выявить основные закономерности парабиоза.
До воздействия парабиотического агента сохраняются соотношения между силой раздражения и силой мышечного сокращения. Сильное раздражение нерва сопровождается сильным мышечным сокращением.
В начальной фазе действия агента несколько уменьшается сила мышечного сокращения и в том и другом случае.
В первой — трансформирующей (уравнительной или провизорной) стадии сила мышечных сокращений при сильных и более слабых раздражителях как бы уравнивается по своей величине. Проводимость нерва в большей степени понижается для сильных раздражителей. Следовательно, в этой фазе заданный ритм волн возбуждения превращается в другой (г.е. трансформируется) ритм. Причина — понижение лабильности нерва. Эта стадия предшествует более выраженным изменениям и поэтому называется еще провизорной.
Электроотрицательность парабиотического участка нерва

Ток во внешней цепи направляется от непораженного участка нерва к парабиоти- ческому (направление тока показано стрелками; парабиотический участок нерва заштрихован).
Во второй — парадоксальной — стадии извращаются соотношения между силой раздражения и силой сокращения мышцы. Сильное раздражение вызывает лишь незначительное начальное сокращение мышцы, а более слабое способно еще вызвать значительное мышечное сокращение, выражающееся в тетанусе. При этом показано, что волны возбуждения, приходящие от непораженного участка нерва, производят тормозящее действие, что проявляется в уменьшении силы мышечного сокращения.
В третьей — тозмозящей — стадии любые возбуждения, достигая пораженного участка нерва, вызываю і только тормозящий эффект. Возбудимость нерва резко снижена. Эта стадия переходит в стадию полного парабиоза.
При исследовании электрических свойств нерва при действии на него патогенных факторов (рис.) удалось показать, что первоначально пораженный участок (в котором развивается парабиоз) приобретает положительный заряд относительно неповрежденных участков (слабо выраженная позитивноегь). Следующий этап характеризуется резкими колебаниями потенциала на уровне или около нулевой линии (0—0). В третьей фазе развивается стойкая и резкая негативность, после достижения которой определенного уровня (критическая линия К—К) развивается полный парабиоз.
Схема изменения электрического состояния нерва при парабиозе
(по Н. Е. Введенскому)

В самые первые стадии своего развития возбуждение в измененном участке подкрепляет слабые волны возбуждения, но умеряет сильные. Поступающий в парабиотический участок импульс и сам парабиотический участок влияют друг на друга — приходящий импульс подкрепляет местное возбуждение парабиотического участка, а последнее сообщает приходящей волне некоторую экзальтацию и затяжной характер. Однако подкрепление возбуждения парабиотического участка приводит к созданию еще большей электроотрицательности, а следовательно, к переводу нерва к более глубокому парабиозу. Но тем, что дальные волны подкрепляют стойкое возбуждение измененного участка и доводят его тем самым до более глубоких стадий парабиоза, они уже сами своим действием на измененный участок начинают преграждать путь следующим себе подобным волнам. Чем более сильная волна, тем больший эффект. Вот от того и получается, что уже в эту первую стадию парабиоза более слабые волны экзальтируются, более сильные умеряются. И в общем получается выравнивание эффектов от волн, проходящих через парабиотический участок к мышце.
В клинической практике с парабиозом приходится сталкиваться в следующих ситуациях:
— при травматических повреждениях (в частности, при наличии сдавливающих нерв рубцов) отчетливо наблюдаются уравнительная и парадоксальная стадии парабиоза. Хирургическое удаление рубца сопровождается (иногда прямо на операционном столе) восстановлением нормальной проводимости нерва;
— рефлекторная контрактура. В основе данного явления, как и в предыдущем случае, лежит возникновение очага патологической импульсации под влиянием тех или иных причин (во вторую фазу парабиоза) в моторной сфере. При усилении патогенного раздражителя импульсное возбуждение может смениться парабиотическим торможением и развитием РЕФЛЕКТОРНОГО ПАРАЛИЧА;
— парабиоз вследствие нарушения кровоснабжения нерва возникает при перемежающейся хромоте, облитерирующем эндартериите, болезни Рейно, контрактуре Фолькмана, ишемическом неврите;
Читайте также:
