
Физиология нервного волокна у животных
Нерв состоит из множества нервных волокон, которые, объединяясь в пучки, образуют нервы. Однако возбуждение по каждому нервному волокну проводится изолированно, не переходя на соседние.
Функциональной единицей нервной системы является нейрон, который представляет собой нервную клетку с отростками. Отростки нервных клеток образуют нервные волокна; они являются проводниками нервных импульсов. Различают мякотные (покрытые миелиновой оболочкой) и безмякотные нервные волокна.
Свойства нервных волокон.Нервные волокна обладают высокой возбудимостью, у мякотных волокон она выше, чем у безмякотных. Лабильность нервных волокон паи высшая. Мякотные волокна могут воспроизводить до 500 импульсов в I с. У безмякотных волокон лабильность очень низкая.
Обмен веществ в нерве очень незначителен. Энергия образуется на 1 г нерва примерно в миллион раз меньше, чем на 1 г мышцы. Нерв практически неутомляем, что обусловлено его низким обменом веществ и высокой лабильностью. Это создает благоприятные условия для выполнения нервом основной функции — проведения импульсов возбуждения (нервных импульсов).
После перерезки нервного волокна его периферический отрезок, отделенный от тела нейрона, дегенерирует. Это свидетельствует о том, что нервная клетка является трофическим центром для нервных волокон, которые не могут существовать вне связи с клеткой.
 |
Рис. 4.24. Схема нервно-мышечного синапса:
1 — неврилемма, переходящая в наружный слой сарколеммы; 2 — осевой ци-
линдр; 3 — митохондрии; 4 — синаптические пузырьки; 5 — миофибрилла,
состоящая из протофибрилл; 6 — синаптическое пространство (щель); 7— пост-
синаптическая мембрана; 8 — пресинаптическая мембрана; 9 — сарколемма
Перерезанные концы центрального отрезка могут регенерировать, они образуют колбы роста — утолщения, которые растут в направлении периферического отрезка. Врастание нервных волокон из центрального отрезка в периферический начинается уже через 2 — 3 дня после перерезки. Регенерация нервных волокон происходит очень медленно (0,3— 1 мм в сутки). Например, в мышцах начальные признаки восстановления функций регенерирующего нерва обнаруживают примерно через 1,5 мес после перерезки. Полная регенерация нервов происходит в течение многих месяцев и даже лет.
Синаптическая передача возбуждения.Передача возбуждения с нервного волокна на мышечное происходит через специализированное структурное образование, обеспечивающее контакт аксона двигательного нейрона с мышечным волокном — нервно-мышечный синапс (рис. 4.24). Он состоит из трех основных элементов: пресинаптической мембраны, синаптического пространства (щели) и постсинаптической мембраны.
Рис. 4.25. Схема рефлек-
торной дуги с обратной
связью:
1 — рецептор; 2 — афферентный нейрон; 3 — про-
межуточный нейрон; 4 — эфферентный нейрон; 5 —
эффектор (мышца); 6 — проприорецептор (мышеч-
ное веретено); 7 — нейрон обратной связи; стрелки —
направление прохождения импульса

Контрольные вопросы
1. Что такое физиологический покой, возбуждение и торможение?
2. Назовите биоэлектрические явления в возбудимых тканях.
3. Охарактеризуйте потенциал покоя и действия, проведение возбуждения по нервному и мышечному волокну.
4. Что такое раздражимость, возбудимость, лабильность?
5. Какова теория мышечного сокращения?
6. Опишите утомление мышц.
7. Каковы свойства нервных волокон?
8. Что такое синаптическая передача возбуждения?
Структурно-функциональная характеристика нервных волокон. Нервные волокна — это отростки нейронов, с помощью которых осуществляется связь между нейронами и с исполнительными клетками. Имеется два типа нервных волокон: миелинизированные и немиелинизированные (безмиелиновые). Оболочку безмиелиновых волокон образуют шванновские клетки (леммоциты), в которые погружаются осевые цилиндры нервных волокон. Оболочку миелинизированных волокон образуют в периферической нервной системе также шванновские клетки (миелоциты, глиальная клетка), формирующие миелин (многослойная обертка мембран миелоцитов — до 100), а в ЦНС — олигодендроциты (глиальные клетки, т.е. те же шванновские клетки). Миелиновая оболочка через равные участки (около 1 мм) прерывается, образуя свободные от миелина небольшие участки — перехваты Ранвье. Основную часть миелина (78% сухого веса) составляют липиды, обеспечивающие изолирующие свойства оболочки. Нервные волокна обеспечивают проведение возбуждения и аксонныйтра транспорт, выполняющий трофическую функцию нейрона.
Выделяют три типа волокон: А, В и С (см. табл. 3).
Таблица 3. Типы волокон в нервах млекопитающих (по Эрлангеру-Гассеру)
| Тип волокон | Диаметр волокна, мкм | Скорость проведения возбуждения, м/с | Длительность абсолютного рефрактерного периода, мс |
| Аα (миел.) | 12-20 | 70-120 | 0,4-1,0 |
| Аβ (миел.) | 5-12 | 30-70 | - |
| Аσ (миел.) | 3-6 | 15-30 | 0.4-1,0 |
| Аγ (миел.) | 2-5 | 12-30 | - |
| В (миел.) | 1-3 | 5-12 | 1,2 |
| С (немиел.) | 0,3-1,3 | 0,5-2,3 |
Волокна типов А и В являются миелинизированными. А-волокна — это афферентные и эфферентные волокна соматической нервной системы; к волокнам типа В принадлежат преганглионарные волокна ВНС. С-волокна немиелинизированные — это постганглионарные волокна ВНС, а также афферентные волокна от некоторых болевых, тепловых и висцеральных рецепторов.
Аксонный транспорт. Основная масса веществ образуется в трофическом центре нейрона, расположенном преимущественно около ядра, а используются эти вещества в различных участках тела нейрона и его отростках. В аксонных окончаниях также происходит синтез медиаторов, АТФ и повторное использование мембраны пузырьков после высвобождение медиатора. Выделяют быстрый и медленный аксонный транспорт (оба они с затратой энергии).
Быстрый аксонный транспорт осуществляется со скоростью 200—400 мм/сут от тела клетки до аксонных окончаний — прямой (антероградный) — и в противоположном направлении — обратный (ретроградный) транспорт. Вещества транспортируются с помощью микротрубочек и микрофиламентов, часть которых представляет собой актиновые нити (актин составляет 10—15 % белков нейрона). Посредством прямого транспорта в аксонные окончания доставляются митохондрии, ферменты, медиаторы, липиды, везикулы, содержащие гликопротеиды мембран, специальные белки и пептиды (нейротрофогены). Посредством обратного транспорта в тело нейрона переносятся везикулы, содержащие остатки разрушенных структур, фрагменты мембран, факторы роста нервов и другие ростовые факторы, регулирующие синтез белка в соме клетки. Многие вещества, доставленные посредством ретроградного транспорта, подвергаются разрушению в лизосомах. В патологических случаях по аксону к телу клетки могут транспортироваться столбнячный экзотоксин, вирусы полиомиелита, герпеса, бешенства.
Медленный аксонный транспорт идет в прямом направлении и представляет собой передвижение всего столба аксоплазмы со скоростью 1—2 мм/сут. С помощью этого транспорта перемещаются образованные в эндоплазматической сети белки микротрубочек и микрофиламентов (актин, тубулин и др.), ферменты цитозоля, РНК, белки каналов, насосов и другие вещества.
Значение аксонного транспорта: 1) необходим для поддержания структуры нервного волокна; 2) необходим для аксонного роста и образования синаптических контактов; 3) играет важную роль при регенерации нервных волокон. На мышечное волокно такое влияние оказывают нейротрофогены (специальные белки, β-эндорфин и другие пептиды); обратное влияние на мотонейрон осуществляется с помощью миотрофогенов (фактора роста нервов, инсулиноподобного фактора роста).
Результат повреждения нервного волокна. Если нервное волокно в результате травмы разорвано, его периферический отрезок, лишенный связи с телом нейрона, подвергается разрушению, которое называется валлеровской дегенерацией. В течение 2-3 сут наступает распад нейрофибрилл, митохондрий, миелина и синаптических окончаний. Участок волокна, связанный с телом нейрона, регенерирует, проводимость восстанавливается. В норме нервное волокно, как и нейрон, функционирует на протяжении всей жизни организма и проводит возбуждение без декремента (затухания) по всей длине волокна, например от тела мотонейронов спинного мозга до мышечных волокон конечностей (до 1 м).
Механизм проведения возбуждения. Проведение ПД возможно только при наличии на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциалзависимых ионных каналов, ответственных за формирование новых ПД. Это осуществляется, согласно нашим представлениям, следующим образом. В распространении ПД можно выделить два этапа: этап распространения электрического поля, снижающего мембранный потенциал, и этап генерации новых ПД в новых участках нервного волокна. Электрическое поле — разновидность материи, посредством которой осуществляется силовое воздействие на электрические заряды, находящиеся в этом поле. Электрическое поле, которое генерируется биологическими структурами, является источником информации о состоянии клеток и органов организма. Например, состояние электрического поля сердца, записанного в виде электрокардиограммы, помогает выявить его возможные повреждения. В зависимости от расположения и концентрации ионных каналов, в мембране нервного или мышечного волокна имеется два варианта проведения ПД: непрерывное и сальтаторное (скачкообразное).
Непрерывное проведение ПД происходит в мышечных волокнах и в безмиелиновых нервных волокнах (тип С), имеющих равномерное распределение потенциалуправляемых ионных каналов по всей длине волокна. Проведение нервного импульса начинается (как и в мышечном волокне) с распространения колеблющегося по величине электрического поля. Амплитуда ПД в нервном волокне (мембранный потенциал + инверсия) составляет 100-120 мВ, постоянная длины мембраны (λm— расстояние, на котором сохраняется 37 % величины ПД в виде электрического поля) в безмиелиновых волокнах равна 0,1—1,0 мм. В связи с этим возникший ПД за счет действия своего электрического поля способен деполяризовать мембрану соседнего участка до критического уровня на расстояние от 0,1 до 1,0 мм. Это означает, что на этом участке (0,1-1,0 мм) одновременно генерируются новые ПД, обусловленные движением ионов Na + в клетку, К + — из клетки (на распространение электрического поля время не затрачивается). Число одновременно возникающих ПД ограничивается длиной возбужденного участка — для безмиелинового волокна 0,1-1,0 мм (ПД возникают рядом друг с другом в непосредственной близости). Причем сами ПД не перемещаются (они исчезают там, где возникают). Главную роль в возникновении новых ПД играет передний ПД. Вспомогательную роль в генерации новых ПД в невозбужденных участках нервного волокна играют промежуточные ПД (возникшие сзади переднего ПД) — их электрическое поле суммируется с электрическим полем переднего ПД, но они находятся дальше от участка нервного волокна, где возникают новые ПД. Таким образом, непрерывное распространение нервного импульса, как и сальтаторное (см. ниже), идет через генерацию новых ПД по эстафете, когда каждый участок мембраны сначала выступает как раздражаемый электрическим полем, а затем — как раздражающий (в результате формирования в нем новых ПД) (рис. 15).
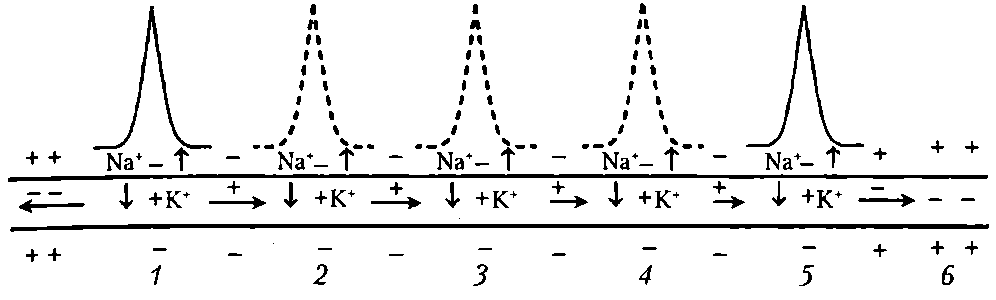
Рис. 15. Непрерывное проведение возбуждения (ПД) в безмиелиновом нервном волокне. Уменьшение длины горизонтальных стрелок иллюстрирует ослабление электрического поля переднего ПД, инициирующего возбуждение соседнего участка волокна: 1-5 - состояние возбуждения (ПД); 6 – состояние покоя; пунктиром обозначены промежуточные ПД; вертикальные стрелки указывают направление движения Na + в клетку и K + - из клетки
Сальтаторное проведение ПД происходит по миелинизированным волокнам (типы А и В), для которых характерна концентрация потенциалуправляемых ионных каналов только в небольших участках мембраны (в перехватах Ранвье), где их плотность достигает 10 тыс. на 1 мкм 2 , что примерно в 100 раз выше, чем в любом участке мембраны безмиелиновых волокон. В области миелиновых муфт, обладающих хорошими изолирующими свойствами, потенциалуправляемых каналов почти нет, поэтому ПД здесь не возникают. ПД, возникший в одном перехвате Ранвье, за счет действия своего электрического поля деполяризует мембрану соседних перехватов до КП, что приводит к возникновению в них новых ПД, т.е. возбуждение возникает как бы скачкообразно — только в перехватах (рис.16).
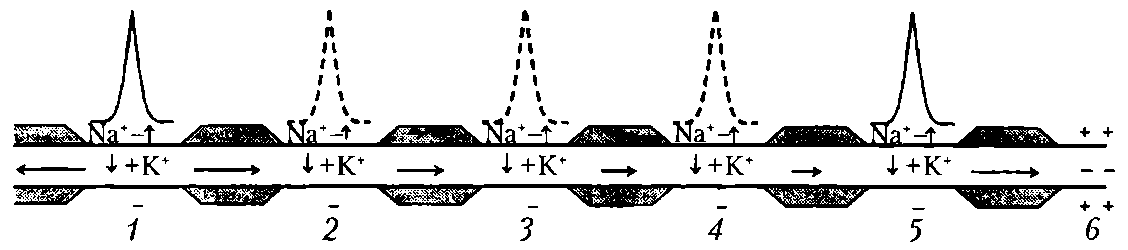
Рис. 16. Сальтаторное проведение возбуждения (ПД) в миелинизированном нервном волокне. Уменьшение длины горизонтальных стрелок иллюстрирует ослабление электрического поля переднего ПД, инициирующего возбуждение соседнего участка волокна: 1-5 - состояние возбуждения (ПД); 6 – состояние покоя; пунктиром обозначены промежуточные ПД; вертикальные стрелки указывают направление движения Na + в клетку и K + - из клетки
Напомним, что Nа-каналы начинают открываться при достижении деполяризации клеточной мембраны 50% КП. Постоянная длины мембраны миелинового волокна достигает 5 мм. Это означает, что электрическое поле ПД на данном расстоянии сохраняет 37% своей амплитуды (около 30 мВ) и может деполяризовать мембрану до КП (ΔV в перехватах Ранвье составляет около 15 мВ). Благодаря этому, в случае повреждения ближайших на пути следования перехватов Ранвье ПД возбуждает 2—4-й и даже 5-й перехваты. Поэтому возбуждение распространяется очень быстро по всей длине волокна, а ионы движутся только перпендикулярно относительно длины волокна — в клетку и из клетки (вдоль волокна они не успевают смещаться). Электрическое поле ПД, возникших сзади переднего (промежуточные ПД), суммируется с электрическим полем переднего ПД, как и при непрерывном распространении возбуждения.
Простые расчеты показывают, что скорость проведения ПД по нервному волокну за счет движения ионов вдоль волокна была бы слишком низкой. В частности, скорость движения Na + в клетку согласно электрохимическому градиенту легко рассчитать по толщине клеточной мембраны (6—10 нм) и длительности ПД (например, толстого миелинизированного волокна — около 1 мс) — вход Na + в клетку и выход К + из клетки. При этом Na + при движении в клетку согласно электрохимическому градиенту преодолевает расстояние в 8 нм примерно за 0,5 мс (восходящая часть пика ПД продолжается около 0,5 мс). На основании этого рассчитаем, сколько времени потребуется на прохождение ПД 1 м. Для прохождения ионом Na + 8 нм необходимо 0,5 мс.
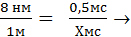
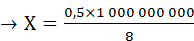
≈17 часов,
т.е. возбуждение распространилось бы на 1 мм за 17 часов.
Легко представить, какова была бы подвижность (точнее, неподвижность) живых существ, если бы возбуждение вдоль нервных и мышечных волокон распространялось со скоростью диффузии ионов, но очень трудно представить, как бы выглядел весь животный мир! Чтобы шевельнуть пальцем пришлось бы ждать около суток!
Возникающие ПД не могут инициировать развитие других ПД в обратном направлении, так как нервное волокно находится еще в рефрактерном состоянии. Это не противоречит тому факту, что раздражение нервного волокна в эксперименте вызывает распространение возбуждения в двух направлениях, поскольку в этом случае участки нервного волокна по обеим сторонам от места раздражения находятся в состоянии покоя. В натуральных же условиях первый ПД, возникший на мембране тела нейрона, инициирует распространение возбуждения только в одном направлении — по аксону к другой клетке.
Сравнение механизма непрерывного и сальтаторного проведения возбуждения показывает, что различие между ними не принципиально. Оно заключается лишь в том, что очередные ПД в безмякотном волокне возникают на более близком расстоянии друг от друга, поскольку ионные каналы расположены в непосредственной близости друг от друга и непрерывно по всей длине нервного волокна. Поэтому такое проведение и назвали непрерывным.
Однако сальтаторное проведение возбуждения имеет два важных преимущества перед непрерывным. Во-первых, сальтаторное проведение более экономично с точки зрения затрат энергии, так как возбуждаются только перехваты Ранвье, площадь которых менее 1% мембраны, и, следовательно, требуется меньше энергии для восстановления трансмембранных градиентов Na + и К + , расходующихся в процессе возникновения ПД (в миелинизированных волокнах при распространении возбуждения теряется ионов в 100 раз меньше, чем в немиелинизированных). Во-вторых, возбуждение в миелинизированных волокнах проводится с большей скоростью (см. таблицу), чем в безмиелиновых волокнах, так как в них электрическое поле ПД распространяется значительно дальше на соседние перехваты Ранвье, поскольку электроизоляция (миелиновые муфты) уменьшает рассеивание электрического поля.
В процессе проведения возбуждения время затрачивается только на перпендикулярное относительно мембраны волокна движение ионов в клетку и из клетки при формировании нового ПД, а влияние электрического поля возникших ПД на соседний участок распространяется вдоль длины волокна мгновенно — время распространения электрического поля практически равно нулю. Скорость распространения возбуждения увеличивается также при большой амплитуде ПД, что является следствием формирования более сильного электрического поля, обеспечивающего критический уровень деполяризации нервного волокна на большем расстоянии и быстрее.
В настоящее же время одни физиологи считают, что возникший ПД электротонически вызывает возбуждение в соседнем участке нервного или мышечного волокна; по мнению других, W. Willis (2004) и R. Klinke (2004), это осуществляется с помощью локальных входящих и выходящих токов ионов и между соседними участками волокна в продольном направлении. Однако электротон — это частичная деполяризация мембраны, еще не активирующая ионные каналы, поэтому в первом случае возбуждение вообще не возникло бы, а во втором случае оно распространялось бы слишком медленно.
Скорость распространения электрического поля хорошо иллюстрируется процессом общения по телефону — собеседники могут находиться на расстоянии нескольких тысяч километров друг от друга, но слова они слышат через такой же промежуток времени, как и при непосредственном разговоре.
Характеристика проведения возбуждения по нервному волокну. 1. Двустороннее проведение возбуждения можно продемонстрировать в эксперименте при нанесении раздражения в любом участке нерва или нервного волокна, при этом возбуждение регистрируется как в одном, так и в другом направлении от места раздражения.
2. Изолированное проведение возбуждения в отдельных волокнах нервного ствола обусловлено тем, что влияние электрического поля ПД соседнего волокна не возбуждает другие волокна нерва вследствие изолирующего эффекта их оболочек и интерстиция. Изолированное проведение импульсов по нервным волокнам обеспечивает точное афферентное и эфферентное влияния функционально разнородных волокон нерва. Однако если одновременно возбуждается значительное количество волокон, то возникает достаточно сильное электрическое поле, способное открыть ворота натриевых каналов соседних и таким образом усилить нервное влияние на эффекторные клетки или нейроны в регуляторных процессах.
3. Бездекрементное (без затухания; лат. decrementum — убывание, уменьшение) проведение по всей длине нервного волокна, так как ПД возникают в каждом участке волокна заново под влиянием предыдущего ПД, и его величина в каждом участке волокна складывается из величины ПП и фазы инверсии ПД.
4.Большая скорость проведения возбуждения (до 120 м/с) в нервных волокнах типа Аα. Для сравнения отметим, что скорость передачи гуморальных влияний ограничена скоростью кровотока — от 0,5 мм/с в капиллярах до 0,25 м/с в аорте (время полного кругооборота крови — около 22 с). Большая скорость распространения ПД обеспечивает быстрое влияние на другие нейроны, рабочие органы, получение обратной информации, играющей важную роль в регуляторных процессах.
Малая утомляемость нервного волокна впервые была показана Н.Е. Введенским (1883): в проводимых опытах нерв сохранял способность к проведению возбуждения в течение 6—8 ч непрерывного раздражения несильными импульсами тока при условии наличия кислорода в окружающей среде и поддержания влажного состояния нерва. Это обусловлено тем, что при проведении ПД по нервным волокнам используется незначительная часть запасов трансмембранных ионных градиентов и, следовательно, нужны небольшие количества АТФ для их восстановления. Расход энергии в нервном волокне на единицу массы в миллион раз меньше, чем в работающей мышце.
Высокая лабильность — нервное волокно может проводить до 500—1000 имп./с.
Возбуждение от нервного волокна передается к другой клетке с помощью синапса.
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
СТРУКТУРНЫЕ ОСОБЕННОСТИ НЕРВНЫХ КЛЕТОК И ВОЛОКОН
Для лучшего понимания механизмов процесса возбуждения в нервных структурах кратко остановимся на их строении.
Нервную систему высших организмов делят на соматическую и автономную. Соматическая участвует в регуляции функционирования внешних покровов и скелетных мышц животных, автономная — в регуляции функций внутренних органов. Оба типа нервной системы имеют центральный и периферические отделы. В центральный отдел входят спинной и головной мозг.
Периферический отдел включает нейроны, сгруппированные в особые образования — ганглии, или же отдельные нейроны, находящиеся вблизи различных структур организма, регуляцию функций которых они осуществляют (см. гл. 12). Как показали многочисленные морфологические исследования, нейроны из различных отделов нервной системы животных могут быть разнообразной формы и размеров. Вместе с тем в строении нейронов можно выделить общие черты. Нейрон состоит из тела (сомы), в котором находится клеточное ядро, и отростков (рис. 2.1). Отростки получили название дендритов и аксонов. Дендриты подводят к соме возникающее в их разветвлениях возбуждение. Аксоны, наоборот, передают от нейрона возбуждение через свои разветвления к другим клеткам. Следует отметить, что название отростков нейрона первоначально дано морфологами на основании предположений о том, что короткие и сильно ветвящиеся отростки, напоминающие по форме ветвление дерева (отсюда и название дендрит), подводят к нейрону возбуждающие сигналы, а длинные маловетвящиеся отростки отводят от сомы нейрона возбуждение. Однако последующие нейроморфологические и нейрофизиологические исследования показали, что по этим критериям невозможно определить функциональное назначение отростков нейрона, поскольку существуют значительные вариации как в степени ветвления, так и длины аксонов и дендритов. Размеры (диаметр) сомы нейронов у сельскохозяйственных животных варьируют от 5 до 100 мкм, а диаметр аксонов и дендритов — в пределах 0,3. 25 мкм.

Рис. 2.1. Различные типы нервных клеток:
I — ядро; 2— сома клетки; стрелками показано направление распространения возбуждения в нервных волокнах
В то же время отростки нервных клеток (аксоны и дендриты), например иннервирующие нижнюю конечность у коровы или лошади, могут быть длиной более 1 м.
Нейроны по количеству отростков, отходящих от сомы, можно разделить на три группы (см. рис. 2.1): униполярные нейроны с одним отростком; биполярные — с двумя отростками; мульти- полярные, имеющие более двух отростков и чаще всего встречающиеся у позвоночных животных. Нейроны, как и клетки других типов, снаружи покрыты плазматической мембраной. Она отделяет цитоплазму клетки от внеклеточной жидкости и выполняет ключевую роль в процессе возбуждения нейрона. Цитоплазма нейрона помимо ядра содержит аппарат Гольджи, систему микротрубочек, нейрофибриллы, митохондрии, рибосомы. С помощью своих отростков нервные клетки контактируют друг с другом, а также с другими типами клеток в организме животного, образуя сложные сплетения.
Контакты между клетками имеют специальное строение и называются синапсами. У позвоночных животных обнаружены различные типы синапсов, различающиеся по структуре и функции. Наиболее часто встречаются так называемые химические

Рис. 2.2. Схематическое изображение миелинизированного нервного волокна:
А, Б и В — различные стадии развития мислиновой оболочки, Г — мислинизиро- ванное нервное волокно, 1 — шванновская клетка; 2 — нервное волокно, 3 — перехват Ранвье, 4— слои миелина
синапсы, в которых передача сигнала осуществляется с помощью специального химического вещества-передатчика — медиатора. К телу нейронов и их отросткам тесно прилегают глиальные клетки (астроциты, олигодендроциты, шванновские клетки и др.). Число глиальных клеток в организме животного превышает количество нейронов более чем в 10 раз. Среди их предполагаемых функций достаточно хорошо изучена функция, связанная с образованием миелиновой оболочки вокруг отростков нейронов.
В морфологических исследованиях с использованием электронного микроскопа было убедительно продемонстрировано, что в периферических нервных окончаниях во время эмбрионального и постнатального развития шванновские клетки многократно обматываются вокруг нервного отростка, формируя цилиндрическую оболочку, состоящую вначале из чередующихся слоев мембраны и тонкого слоя цитоплазмы (рис. 2.2). Затем слой цитоплазмы исчезает и образуется компактная структура, содержащая в основном липиды, получившая название миелиновой оболочки. Одна шванновская клетка может покрыть нервное волокно миелиновой оболочкой на расстоянии 0,9. 2 мм. Мембрана нервного волокна остается неприкрытой на узком промежутке длиной около 1 мкм. Этот участок нервного волокна получил название перехвата Ранвье.
Читайте также:
