
Гены развития нервной системы
Впервые ранние гены были идентифицированы в культуре ткани при исследовании механизмов активации генома в ответ на добавление в среду факторов роста, приводящих к клеточной пролиферации (увеличению массы клеток путем митоза) и дифференцировке. Эти исследования привели к открытию класса генов, которые активируются в течение нескольких минут после воздействия на клетки факторов роста, причем эта транскрипционная индукция происходит даже в том случае, если в среду добавлены ингибиторы синтеза белка. Оказалось, что эти гены кодируют ядерные белки, которые являются транскрипционными факторами и регулируют экспрессию других генов.
Одними из первых в семействе ранних генов были идентифицированы гены c-fos и с-тус.
Оказалось, что ранние гены широко распространены в живых организмах, причем один и тот же ген встречается у далеких в эволюционном отношении организмов. Это указывает на их важную биологическую функцию в нормальных клетках. Было показано, что ранние гены в условиях нормального функционирования характеризуются быстрой, но преходящей экспрессией в ответ на различные сигналы из внеклеточного пространства. Они способны реагировать на нейротрансмиттеры (передатчики нервных импульсов), на различные белковые факторы роста (например, фактор роста нервов - см. далее). Таким образом, действие разнообразных экстраклеточных сигналов приводит к запуску в клетках систем вторичных мессенджеров (посредников) и активации различных транскрипционных факторов, способных взаимодействовать друг с другом и активировать синтез мРНК генов-мишеней, обеспечивая тем самым специфичность и разнообразие восприятия клеткой различных сигналов.
Считается, что ранние гены и их продукты могут быть одним из механизмов, посредством которых осуществляется координация между онтогенетическими событиями в нервной системе и во внешней среде. Посредством участия ранних генов некоторая генетическая активность может концентрироваться в определенных клетках и координироваться с другими процессами, происходящими в нервной системе или во внешней среде.
Исследования показывают, что ранний опыт может значительно влиять на экспрессию ранних генов в клетках коры головного мозга. Так, К.М. Розен (1992) измерял уровень содержания мРНК ранних генов в зрительной и фронтальной коре котят, воспитывавшихся в темноте. Оказалось, что даже короткая световая экспозиция (всего один час) драматически увеличивает активность ранних генов в зрительной, но не во фронтальной коре. Примечательно, что эта активность была выше, если световая экспозиция приходилась на более ранний период развития (5-недельные котята в сравнении с 20-недельными). Таким образом, опыт может регулировать молекулярную генетическую активность. Накопленные данные о механизмах экспрессии ранних генов позволили предположить, что они могут не только играть важную роль в развитии нервной системы, но и быть связанными с такими важными процессами, как обучение и консолидация памяти (то есть процесс перехода информации из кратковременной памяти в долговременную). К настоящему моменту накоплено очень много данных об экспрессии ранних генов в мозге животных при обучении (см. например: Анохин К.В., Судаков К.В., 1993). Обобщая полученные результаты, можно выделить несколько основных свойств экспрессии ранних генов в мозге при обучении.
В обычном состоянии, при отсутствии во внешней среде значимых раздражителей, экспрессия ранних генов в мозге животных осуществляется на базальном уровне с очень слабой интенсивностью. При помещении животных в новую среду или обучении их новым навыкам происходит очень быстрое и значительное усиление экспрессии ранних генов, причем наблюдаемая индукция транскрипции достигает своего максимума через 15-30 минут после начала воздействия, а уже через 60-90 минут уровень мРНК ранних генов возвращается к первоначальному базовому значению. Соответственно максимальный синтез белков, кодируемых этими генами, детектируется через 1-2 часа после воздействия, и через 4-5 часов индукция завершается (Там же).
Примечателен тот факт, что по мере выработки и автоматизации нового навыка уровень экспрессии генов c-fos и c-jun постепенно снижается, а выполнение уже приобретенного навыка не вызывает индукции ранних генов. Привыкание к процедуре, вызывающей у животного стресс, также приводит к значительному снижению экспрессии ранних генов. При замене некоторых объектов во внешней среде или частичном изменении некоторых параметров условного сигнала затухавшая по мере обучения экспрессия ранних генов вновь достигает почти первоначального уровня (Там же).
Эти данные послужили основой для гипотезы о том, что активация в мозге ранних генов происходит в том случае, когда животное сталкивается с новыми для него ситуациями и возникающая при этом экспрессия ранних генов отражает субъективную оценку животным новизны (Там же).
В последние годы были получены прямые доказательства участия ранних генов в механизмах обучения и формирования памяти. Современные методы молекулярной биологии позволяют специфично и обратимо блокировать экспрессию отдельных генов. Этого можно добиться, вводя в клетки короткие последовательности олигонуклеотидов, комплементарных участку мРНК интересующего гена. Связавшись с мРНК, такие антисмысловые (антисенс-) последовательности блокируют трансляцию, и в клетке не синтезируется белок данного гена. В экспериментах на крысах и цыплятах было показано, что введение в критические для запоминания области мозга антисенс-олиго-нуклеотидов, временно "выключающих" экспрессию гена c-fos, вызывает у обученных животных амнезию.
Эти данные свидетельствуют о том, что ранние гены очень чувствительно реагируют на изменения во внешней среде и являют собой критическое звено в механизме воздействия среды и приобретаемого в результате этого индивидуального опыта над геном нервных клеток.
Таким образом, благодаря ранним генам даже краткие средовые сигналы могут быть преобразованы в каскад продолжительных событий, приводящих к изменениям внутри клеток и к межклеточным взаимодействиям. Тем самым ранние гены могут участвовать как в закладке нервной системы в эмбриогенезе, так и в ее пластических изменениях после рождения.
Помимо ранних генов, в регуляцию генной активности и межклеточного взаимодействия включены и другие продукты. Одними из них являются гормоны.
Короткие белки, контролирующие работу нейроэндокриноиммунной системы и ее центральных отделов, называют нейропептиды. Некоторые белки являются гормонами (нейрогормонами). Их наличие особенно ярко демонстрирует участие генов в формировании поведенческих признаков, включая характеристики психической активности мозга человека. Так, широко изучается роль нейропептидов галанина и NPY, а также белка лептина в регуляции пищевого поведения. Формирование материнского поведения у самок крыс регулируется изменением содержания нейропептида тахикинина в вентромедиальном ядре гипоталамуса. Кортикотропин-рилизинг фактор гипоталамуса определяет эмоциональность в реакциях животных на новизну. Гормон гипофиза вазопрессин, как и его аналог AVT (аргинин-вазопрессин) у птиц, влияет на территориальное поведение животных.
| Табл. 6. Некоторые белковые молекулы, контролирующие процессы роста нейронов, узнавания нейронами друг друга и синаптической передачи | ||
| Нейротрофные факторы | Молекулы распознавания | Пре- и постсинаптические молекулы |
| NGF, р75, Trk A, GDNF, BDNF, NT 3, 4 и 5 CNTF, ILGF I и II FGF EGF и др. | PSA-NCAM NCAM (H-CAM, G-CAM, VCAM-1 и др.) APP, Интегрин Ламинин, Кадгерин Тенцин, протеогликаны и др. | ά-GDI, Rab3A RIM, Munc13 Синаптотагмин Олигофренин G-белок PAK-3 и др. |
Следует понимать, что на формирование нервной системы могут негативно влиять и нарушения в генах, работающих в клетках других тканей. Косвенно эти нарушения могут отражаться на работе нейроэндокриноиммунной системы. Чем раньше в процессе становления нервной системы проявляется дефект какого-либо гена, тем сильнее это сказывается на развитии организма и его последующем поведении. Внешние стимулы также регулируют работу генов уже на ранних стадиях развития эмбриона. Показано, что действие стрессоров на пренатальных стадиях развития у лабораторных крыс сказывается на структуре хроматина ядер их нервных клеток. Это отражается на режиме работы генов в клетках-мишенях. Измененная структура хроматина сохраняется в нервных клетках уже родившихся животных. Впоследствии, спустя даже длительный период времени, их поведение отличается от нормального.
Определение:
Хроматин– специфически окрашиваемые области ядра клетки, в которых расположена ДНК и связанные с ней белковые компоненты.
Определение:
Трансгенныеживотные – животные, в геном которых была встроена чужеродная (т.е. не принадлежащая данному животному) ДНК.
Говоря о различных свойствах человеческой психики (будь то темперамент, интеллект, экстра-интроверсия и т.д.) большинство современных исследователей признают их (свойств) зависимость от физиологических, эндокринных и биохимических детерминант (Малых и др., 1998). Остается сделать еще один шаг и сказать, что все вышеперечисленные биологические детерминанты в той или иной степени контролируются генетически. Т.о., одной из задач современной психогенетики является поиск генетических механизмов контроля этих детерминант, т.е. фактически генетического контроля психики. При этом речь идет не только о генетической зависимости типовых психических особенностей высокоразвитых животных и человека, но и о конкретных индивидуальных различиях.
На современном этапе продолжается активное проникновение нейрофизиологии и нейрогенетики в психологию. Изучаются физиолого-генетические механизмы реализации психологического фенотипа. Отталкиваясь от работ И.П. Павлова о типах высшей нервной деятельности, а также от представлений В. Вундта, Г. Айзенка и других, исследователи развивают генетические представления о наследуемости свойств нервной системы. Изучаются гены, влияющие на эмоциональность, социабильность, заторможенность и активность. Анализируется роль нейропептидов в регуляции сложно организованных форм поведения, ведется поиск генов, участвующих в синтезе нейромедиаторов и нейрогормонов. Совершенствуются представления о сети генов единой нейроэндокриноиммунной системы (НЭИС) высших животных как механизма, обеспечивающего максимально быстрый адаптивно значимый ответ организма на любое изменение в окружающей среде. Перспективными моделями в этой связи представляются изучение генетического контроля стресс-реакции организма, а также анализ формирования НЭИС в развитии организма и определение чувствительных стадий развития. Генетика нейрофизиологических признаков организма лежит в основе понимания закономерностей становления психогенетических признаков животных и психической активности человека как высшей формы адаптивного поведения.
Задания для самоконтроля:
1. Расскажите о генетическом контроле процесса становления нервной системы в онтогенезе.
4. Расскажите о применении трансгенных животных в изучении функционирования центральной нервной системы.
6. Что такое нейропептиды? Приведите конкретные примеры их наследственной обусловленности и действия в центральной неврной системе.
Семинар 1.
Учебные вопросы:
1. Требования к модельным объектам.
2. Современные молекулярно-генетические методы исследования Ц.Н.С.
3. Механизмы формирования кратковременной и долговременной памяти.
Темы докладов:
- Взгляды Рамон-и-Кахаля на природу индивидуальности человека и их развитие в современных молекулярно-генетическиех исследованиях.
- Современные представления о процессах запоминания.
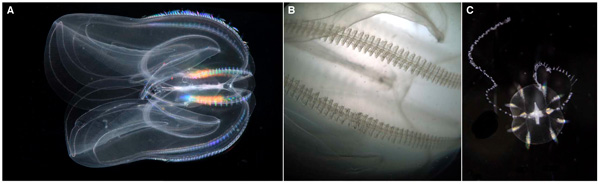

Исследование недавно прочитанного полного генома гребневика Mnemiopsis leidyi привело к выводу, что тип гребневиков является самой древней ветвью многоклеточных животных, даже более древней, чем губки. Такое положение гребневиков на эволюционном древе значительно подкрепляет гипотезу, согласно которой они приобрели нервную систему независимо от всех остальных. Иными словами, получается, что нервная система возникла дважды.
Однако дать на этот вопрос окончательный ответ очень долго не удавалось. Собственно говоря, такого ответа нет и сейчас. Палеонтологическая летопись дает о гребневиках относительно мало информации: тело этих животных очень нежное, и они плохо сохраняются в ископаемом состоянии. Анатомия гребневиков изучена прекрасно; она очень интересна, но основанные на ней гипотезы о родстве оказались противоречивыми, и ни одна из них так и не стала общепризнанной. Значит, остается молекулярная филогенетика, то есть анализ нуклеотидных последовательностей. Для такой сложной группы его лучше делать сразу по большому числу генов, а в идеале — по всему геному.

Рис. 2. Проблема положения гребневиков на эволюционном древе животных. Cnidaria — кишечнополостные, Bilateria — двусторонне-симметричные животные, Placozoa — пластинчатые, Porifera — губки, Choanoflagellata — воротничковые жгутиконосцы, Ctenophora — гребневики. Ветвь гребневиков помещена в стороне. Здесь показано четыре варианта ее расположения на древе (оранжевые стрелочки); все они соответствуют гипотезам, поддержанным кем-то из зоологов. Гипотеза 1 означает, что гребневики являются ближайшими родственниками кишечнополостных; гипотеза 2 — что они родственники двусторонне-симметричных животных; гипотеза 3 — что они являются самой древней ветвью животных, не считая губок; и гипотеза 4 означает, что гребневики — это самые древние многоклеточные животные вообще (возможно, не считая некоторых ископаемых). Именно гипотезу 4 подтверждают результаты исследования полных геномов. Схема из обсуждаемой статьи в Science, с изменениями
Совсем недавно появилась статья большой группы американских исследователей с анализом полного генома гребневика Mnemiopsis leidyi (рис. 1). Этот геном относительно маленький: в нем примерно 150 миллионов пар нуклеотидов (у человека, например, более 3 миллиардов). Для его сравнения с геномами других животных были параллельно применены два разных вычислительных метода: метод максимального сходства (см.: Maximum likelihood) и байесовский анализ (см.: Bayesian inference in phylogeny). Результаты получились запутанными. Часть расчетов приводила к выводу, что гребневики — самая древняя ветвь животных (среди вообще всех многоклеточных животных, гипотеза 4 на рис. 2), а часть почему-то указывала на их родство с губками. Последнее не подтверждается никакими морфологическими данными, а потому выглядит сомнительно. Для разрешения этого конфликта была построена большая серия деревьев с использованием разных баз данных, разных методик и разных выборок исследованных организмов, и из нескольких возможных гипотез самую большую поддержку (5 деревьев из 16) получила все-таки гипотеза, согласно которой гребневики — сестринская группа всех остальных животных без исключения (рис. 3).
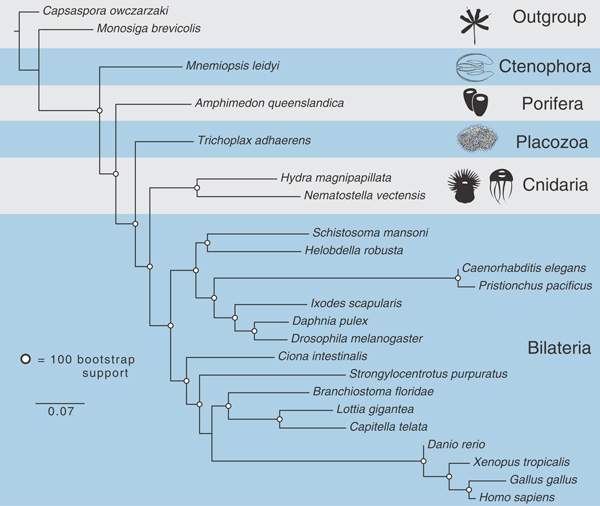
Рис. 3. Одно из итоговых эволюционных деревьев, помещенных в обсуждаемой статье. Значение латинских названий то же, что и на рис. 2; outgroup — внешняя группа, включающая одноклеточных. Длина ветвей соответствует числу нуклеотидных замен. Белыми кружочками отмечены узлы древа, получившие 100-процентную бутстрэп-поддержку. Иллюстрация из обсуждаемой статьи в Science
А вот гипотеза о родстве гребневиков с кишечнополостными была отвергнута всеми расчетами начисто. Так что сходные признаки гребневиков и медуз или приобретены независимо (конвергенция), или, возможно, в какой-то степени унаследованы от общего предка. Но как же этот предок мог выглядеть.
Если гипотеза американских авторов верна, то общая картина ранней эволюции многоклеточных животных видится примерно так. Прежде всего, от общих предков отделилась ветвь, ведущая к гребневикам. Потом — ветвь губок. А уже дальше пошла ветвь, ведущая к кишечнополостным и двусторонне-симметричным (включая нас). Такая эволюционная схема порождает много вопросов, самый актуальный из которых: когда и как возникла нервная система?
У кишечнополостных, как и у двусторонне-симметричных, нервная система есть. У гребневиков — тоже, и не зачаточная, а довольно сложная. А вот у губок, которые вроде бы отделились от общего ствола позже гребневиков, никакой нервной системы нет. И это приводит к очень интересному предположению. Возможно, что нервная система у животных возникала дважды: отдельно у гребневиков и у всех остальных (кишечнополостные + двусторонне-симметричные).
Факты, говорящие за эту идею:
-
Нервная система гребневиков представляет собой дифференцированную сеть, включающую нервные узлы, связанные с органами чувств и основаниями щупалец. Эти структуры не имеют близких аналогов ни у каких других животных, даже и у кишечнополостных (которые тоже относительно примитивны и похожи на гребневиков по типу симметрии).
Надо заметить, что гипотеза о независимом происхождении нервной системы гребневиков высказывается уже не впервые (см.: E. Pennisi. 2013. Nervous System May Have Evolved Twice). Похоже, что она становится популярной.
В итоге речь идет сразу о трех взаимосвязанных гипотезах:
Эти гипотезы могут быть верны одновременно, но, в принципе, могут и по отдельности. Пока еще ни одна из них не подтверждена настолько достоверно, чтобы быть включенной в учебники. Но задуматься они, конечно, заставляют.
Источник: Joseph F. Ryan, Kevin Pang, Christine E. Schnitzler, Anh-Dao Nguyen, R. Travis Moreland, David K. Simmons, Bernard J. Koch, Warren R. Francis, Paul Havlak, NISC Comparative Sequencing Program, Stephen A. Smith, Nicholas H. Putnam, Steven H. D. Haddock, Casey W. Dunn, Tyra G. Wolfsberg, James C. Mullikin, Mark Q. Martindale, Andreas D. Baxevanis. The Genome of the Ctenophore Mnemiopsis leidyi and Its Implications for Cell Type Evolution // Science. 13 December 2013. V. 342. P. 1336. DOI: 10.1126/science.1242592.
Аннотация научной статьи по ветеринарным наукам, автор научной работы — Даев Е.В.
В работе кратко обсуждается связь различных стрессоров с работой генома клеток стрессируемого организма. Нарушения стабильности и целостности геномов при стресс-реакции могут рассматриваться не только как следствие, но и как причина разнообразных отклонений и расстройств работы человеческого организма, подвергающих опасности его здоровье
Похожие темы научных работ по ветеринарным наукам , автор научной работы — Даев Е.В.
ИЗБРАННЫЕ ВОПРОСЫ БИОЛОГИИ И МЕДИЦИНЫ
СТРЕСС, НЕРВНАЯ СИСТЕМА, ГЕНЫ И ЗДОРОВЬЕ
2ФГБУН Институт физиологии им. И.П. Павлова РАН, Санкт-Петербург
В работе кратко обсуждается связь различных стрессоров с работой генома клеток стрессируемого организма. Нарушения стабильности и целостности геномов при стресс-реакции могут рассматриваться не только как следствие, но и как причина разнообразных отклонений и расстройств работы человеческого организма, подвергающих опасности его здоровье. Ключевые слова: стресс, нервная система, гены, здоровье
E.V. Daev 1 2, N.A. Dyuzhikova2 Stress, nervous system, genes and health. The paper briefly discusses the relationship of various stressors to the work of the genome of cells of a stressed organism. Violations of the stability and integrity of genomes under stress reactions can be considered not only as a consequence, but also as a cause of various deviations and disorders of the human body, endangering its health. Key words: stress, nervous system, genes, health
Нервная система (НС) любого многоклеточного организма - это механизм, обеспечивающий связь клеток этого организма с окружающей средой. Именно чувствительность НС к малейшим изменениям в окружающей среде, позволяет всем задействованным в ответе клеткам организма адаптироваться, в основном за счет модуляции работы генетического материала, изменений клеточного транскриптома и протеома. Адаптация клеток организма к изменившимся условиям и обуславливает его нормальное
функционирование (другими словами - здоровье).
Как известно, сложный многоклеточный организм состоит из огромного числа клеток разных типов. И у каждого типа клеток, у каждой клетки есть свои пределы адаптационных возможностей, за которыми клетка не может нормально существовать. Эти пределы определяются её генотипом и функциональным состоянием, а также взаимодействием с соседними клетками. И если клетка не в состоянии адаптироваться к изменившимся условиям, её геном начинает перестраиваться, и даже разрушаться. Если число подобных клеток в составе организма достаточно велико, это приводит к недостаточности или отказу работы какой-то системы (или органа). Организм заболевает и, в конце концов, погибает [1].
Результатом является дистресс, характерными признаками которого являются в первую очередь нарушения работы собственно НС, иммуносупрессия и подавление репродуктивной функции.
Особую и абсолютно неисследованную проблему представляет дестабилизация при стрессе генома нейронов и клеток глии, что может являться маркером развивающегося в организме стресса и, одновременно, нарушать работу тонких механизмов обработки информации в ЦНС.
Для клеток нервной системы в качестве показателей повреждения ДНК используют однонитевые и двунитевые разрывы. Эти типы повреждений ДНК индуцируются генотоксическими соединениями и могут влиять на процесс транскрипции, либо репликацию и репарацию, приводя к образованию генных мутаций [6]. Двойные разрывы ДНК особенно вредны, поскольку могут приводить к нейродегенерации [7] и апоптозу [8]. Одним из наиболее исследованных маркеров формирования двойных разрывов ДНК является гистон Н2АХ, фосфорилированный по серину 139 (уН2АХ) [9].
Известно, что такие стрессорные для животных воздействия, как вынужденное плавание и иммобилизация, а также сочетанное действие стрессоров вызывают образование двунитевых разрывов ДНК в нейронах [10,11]. Активация нейронов, вызванная стрессом, может приводить к различным результатам, поскольку гормоны стресса замедляют восстановление ДНК [12] и способствуют накоплению повреждений ДНК [13]. В очень немногих работах был исследован постстрессорный уровень двойных разрывов ДНК, причем длительность их сохранения не рассматривалась [14, 15]. В модельных экспериментах на мышах продемонстрировано увеличение уровня уН2АХ в нейронах гиппокампа после стресса; однако механизмы этих изменений остаются пока мало исследованными [11]. Известно, что вызванное стрессом высвобождение адреналина ведет к увеличению уровня двойных разрывов ДНК после повторной процедуры стрессорного воздействия [14].
Вместе с тем возникновение двунитевых разрывов ДНК связано и с проявлением нормальной нейрональной активности, что продемонстрировано в нейронах коры и гиппокампа мышей, активированных новым опытом [16]. При этом уН2АХ быстро возвращается к исходному уровню.
Механизмы быстрого и точного восстановления - репарации имеют решающее значение для сохранения жизнеспособности нейронов, но насколько эффективно они работают при стрессе — неизвестно. Следует отметить, что в клетках зрелой нервной системы действует иной механизм поддержания
стабильности генома, чем в процессе развития (при нейрогенезе), в силу отсутствия клеточных делений, репликативных повреждений ДНК и гомологичной рекомбинации. В некоторых случаях репарация повреждений ДНК неэффективна, может осуществляться с ошибками и приводить к возникновению мутаций. Кроме того, процесс репарации двунитевых разрывов ДНК может сопровождаться эпигенетическими изменениями - метилированием ДНК с последующей репрессией генов. Дестабилизация работы геномов (и даже их дезинтеграция) может идти специфично по отношению к типам клеток центральной нервной системы и тем или иным зонам мозга, а также зависеть от вида стрессорного воздействия и генотипа особи [17]. Необходимо проведение дальнейших исследований для расшифровки механизмов формирования и эффективного восстановления постстрессорных повреждений ДНК в клетках мозга.
Таким образом, социально-индуцированные стрессы - один из мощных источников повышения скорости старения, смертности и снижения рождаемости в современном обществе [18]. Необходима борьба с источниками подобных стрессов путём:
адекватного воспитания с детского возраста;
повышения уровня естественно-научной и духовно-нравственной образованности;
контроля и борьбы с непрофессионализмом, модой и коммерциализацией в науке и средствах массовой информации;
контроля качества социально-значимой информации, предоставляемой широким слоям населения.
Это - залог повышения уровня здоровья населения любой страны.
Исследование поддержано средствами гранта РФФИ № 16-04-00678.
1. Daev EV. Genetic effects of olfactory stress: studies on a house mouse. Saarbrucken: Lambert Academic Publishing; 2011. (In Russ.) (ISBN-13: 978-38443-5969-5; ISBN-10: 3844359699; EAN: 9783844359695)
2. Daev EV, Zabarin AV, Barkova SM, Dukel'skaya AV. Distortions of Scientific Information as a Source of the Formation of Tension in Society: the GMO Case // Russian Journal of Genetics: Applied Research, 2016, Vol. 6, No. 6, pp. 633645. DOI: 10.1134/S2079059716060034
3. Cooper MA, Clinard CN, Morrison KE. Neurobiological mechanisms
supporting experience-dependent resistance to social stress // Neuroscience. 2015. V.291. P.1-14. DOI: 10.1016/j.neuroscience.2015.01.072
4. Tung J, ■ Gilad Y. Social environmental effects on gene regulation // Cell. Mol. Life Sci. 2013. V. 70. P. 4323-4339. DOI: 10.1007/s00018-013-1357-6
5. Muehsam D, Lutgendorf S, Mills PJ., et al. The embodied mind: A review on functional genomic and neurological correlates of mind-body therapies // Neuroscience and Biobehavioral Reviews, 2017. V.73. P.165-181. DOI: 10.1016/j.neubiorev.2016.12.027
6. Vijg J, Uitterlinden AG. A search for DNA alterations in the aging mammalian genome: An experimental strategy // Mech Ageing Dev. 1987. V.41(1-2). P. 47-63. DOI: 10.1016/0047-6374(87)90053-4.
7. Madabhushi R, Pan L, Tsai LH. DNA damage and its links to neurodegeneration// Neuron. 2014. V.83. P. 266-282.
8. Jackson SP. Sensing and repairing DNA double-strand breaks//Carcinogenesis. 2002. V.23. P.687-696
9. Rogakou EP, Pilch DR, Orr AH, Ivanova VS, Bonner WM. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem.1998. V.273. P. 5858-5868.
10. Consiglio AR, Ramos AL, Henriques JA, Picada JN. DNA brain damage after stress in rats // Prog neuropsychopharmacol Biol Psychiatry. 2010. V.34(4). P. 652656. DOI: 10.1016/j.pnpbp.2010.03.004.
11. Hare BD, Thornton TM, Rincon M, Golijanin B, King SB, Jaworski DM, Falls WA. Two Weeks of Variable Stress Increases Gamma-H2AX Levels in the Mouse Bed Nucleus of the Stria Terminalis //Neuroscience. 2018; 373: 137-144.
12. Flint MS, Baum A, Chambers WH, Jenkins FJ. Induction of DNA damage , alteration of DNA repair and transcriptional activation by stress hormones// Psychoneuroendocrinology. 2007. V.32. P.470-479.
13. Hara MR , Kovacs JJ, Whalen EJ, Rajagopal S, Strachan RT, Grant W, Towers AJ, Williams B, Lam CM, Xiao K, Shenoy SK, Gregory SG, Ahn S, Duckett DR, Lefkowitz RJ. A stress response pathway regulates DNA damage through beta2-adrenoreceptors and beta-arrestin-1// Nature , 2011. V.477. P. 349-353.
14. Hara MR, Sachs BD, Caron MG, Lefkowitz RJ. Pharmacological blockade of a beta(2)AR-beta-arrestin-1 signaling cascade prevents the accumulation of DNA damage in a behavioral stress model// Cell Cycle. 2013. V.12. P.219-224.
15. He ZY, Wang WY, Hu WY Yang L, Zhang WY, Yang YS, Liu SC, Zhang FL, Mei R, Xing D, Xiao ZC, Zhang M. Gamma-H2AX upregulation caused by Wip
1 deficiency increases depression-related cellular senescence in hippocampus.// Sci. Rep. 2016, 6: 34558.
16. Suberbielle E, Sanchez PE, Kravitz AV, et al. Physiologic brain activity causes DNA double-strand breaks in neurons, with exacerbation by amyloid-beta. Nat neurosci. 2013;16(5):613-621. DOI: 10.1038/nn.3356. 165.
17. Dyuzhikova NA, Daev EV. Genome and stress-reaction in animals and humans // Ecological genetics. 2018. V. 16(1). P. 4-26. DOI: 10.17816/ecogen1614-26
18. Franzke B, Neubauer O, Wagner K.-H. Super DNAging—New insights into DNA integrity, genome stability and telomeres in the oldest old // Mutat. Res. 2015. V. 766. P. 48-57. DOI: 10.1016/j.mrrev.2015.08.001
Ананьев В.Н.,1 Ананьев Г.В.2
ВЛИЯНИЕ РАБОТАЮЩИХ ПОПЕРЕЧНОПОЛОСАТЫХ МЫШЦ НА АДРЕНОРЕЦЕПТОРЫ АРТЕРИЙ ДО И ПОСЛЕ АДАПТАЦИИ К ХОЛОДУ
1ГНЦРФ Институт медико-биологических проблем РАН, г. Москва, e-mail: noradrenalin1952@mail.ru; 2МПГУ - Московский педагогический государственный университет, г. Москва
Аннотация. В работе представлены механизмы регуляции при моделировании рабочей гиперемии в поперечнополосатой мускулатуре у животных (кроликов) при адаптации к низким температурам в течение тридцати суток. При искусственном кровоснабжении задней конечности животного через насос постоянного расхода оценивалась величина изменения давления в arterial femoralis при введении в артерию восьми возрастающих доз норадреналина и адреналина. По величинам доза-эффект, в двойных обратных координатах Лайниувера-Берка, оценивали реактивность артериальных адренорецепторов. Для моделирования работы мускулатуры и для достижения мышечной рабочей гиперемии применялась электромиостимуляция мускулатуры бедра и голени лабораторного животного. Проведенные исследования показали, что механизм мышечной гиперемии заключается в уменьшении чувствительности адренорецепторов артерий к адреналину и норадреналину при неизменном количестве активных адренорецепторов. При
Читайте также:
