
Из чего состоит нервно мышечный препарат
По отношению к внешнему раздражению ткани организма делятся на невозбудимые (эпителиальная, соединительная, костная) и возбудимые (нервная и мышечная). При раздражении нервной и мышечной ткани в ней возникает возбуждение, которое распространяется по этой ткани вдоль от места раздражения. Нервно-мышечный препарат лягушки является классическим объектом для изучения функций возбудимых тканей.
Цель работы.Научиться выделять возбудимую ткань лягушки для изучения ее свойств.
Оснащение. Набор хирургических инструментов, физиологический раствор для холоднокровных животных, марля, эфир. Работу выполняют на лягушке.
Ход препарирования. Наркотизированную лягушку оборачивают марлей так, чтобы лапки ее были прижаты к туловищу, а голова оставалась свободной. Ножницами отсекают верхнюю челюсть за глазными буграми. В центральный канал вводят зонд и разрушают спинной мозг. При втором способе обездвиживания лягушки большим пальцем наклоняют вперед голову лягушки и находят углубление кзади от затылочной кости. Вертикально вводят в субокципитальное отверстие конец зонда на глубину 1-2 мм, поворачивают зонд параллельно спинномозговому каналу, вводят в него зонд, который продвигают до крестцово-копчикового соединения. Круговыми движениями разрушают спинной мозг. Выводят иглу из центрального канала и, повернув ее на 180 0 , разрушают головной мозг. Критерием разрушения мозга является полное расслабление скелетных мышц лягушки и отсутствие защитных двигательных рефлексов на пощипывание кожи и потягивание за лапку.
Взяв лягушку за задние лапки, поворачивают вниз брюшком и, отступив на 1,5 см выше копчика, перерезают позвоночный столб с окружающими тканями (рис 1). Отделяют внутренние органы и брюшную стенку от задней части туловища. В руке остаются задние лапки с тазовой костью и небольшим отделом позвоночного столба. Захватив край кожи пинцетом, снимают ее с лапок. Вырезают копчик. По средней линии осторожно разделяют лапки, перерезая лобковое соединение. Одну лапку заворачивают в марлю, смоченную физиологическим раствором, с целью сохранения ее для второго эксперимента. На другой лапке подводят лезвие ножниц под пояснично-крестцовое сплетение и отделяют тазовую кость так, чтобы сплетение осталось соединенным с позвоночным столбом. Располагают препарат дорсальной поверхностью вверх. Раздвинув стеклянным крючком двуглавую и полуперепончатую мышцы, находят на бедре седалищный нерв. Приподнимают его и на всём протяжении осторожно отделяют от окружающих тканей. Бедренную кость и мышцы бедра отделяют от препарата выше коленного сустава.
Затем переходят к выделению икроножной мышцы. Для этого отделяют мышцу от голени. Кости голени и другие мышцы голени удаляют, перерезая их ниже коленного сустава и выше пяточного сустава. Стопу отрезают ниже пяточного сустава.
В результате получают нервно-мышечный препарат, который является универсальным объектом для исследования функций скелетных мышц и нервов.
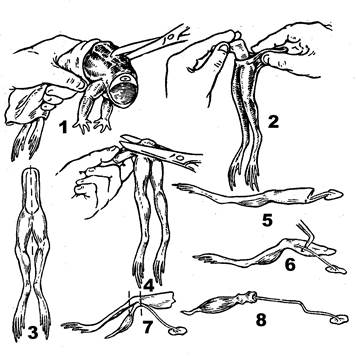 | 1 - перерезка позвоночника после обездвиживания лягушки. 2 - снятие кожи с задних конечностей. 3 - линия разделения нижней части туловища. 4 - удаление копчиковой кости. 5,6 - этапы выделения седалищного нерва. 7 - выделение икроножной мышцы и места перерезки бедра и голени. 8 - нервно-мышечный препарат. |
Рис. 1 Этапы приготовления нервно-мышечного препарата.
Следует помнить, что при небрежном приготовлении препарата (прикосновении к нерву руками или металлическими предметами, натяжении, высыхании, механических повреждениях) препарат быстро теряет возбудимость. Поэтому во время проведения опыта необходимо прикасаться к нерву стеклянным крючком и часто увлажнять нерв и мышцы физиологическим раствором.
Рекомендации к оформлению протокола.
1. Записать ход работы.
2. Зарисовать основные этапы приготовления нервно-мышечного препарата.
Наиболее удобный объект для изучения возбудимости, лабильности и проявлений возбуждения — нервно-мышечный препарат. Нервно-мышечный препарат представляет собой свежепрепарированный из тела лягушки седалищный нерв в соединении с икроножной мышцей. Раздражение нерва происходит в том месте, где прилагается раздражитель. Если оно достаточно сильно, то в этом месте возникает возбуждение, которое проводится нервом к мышце и вызывает ее сокращение. Это позволяет установить возникновение возбуждения в нерве без всяких приборов, только по сокращению мышцы. Вместе с тем к нервно-мышечному препарату очень легко присоединить специальные регистрирующие приборы и детально изучить изменения возбудимости, лабильности и проявления возбуждения. Это изучение можно проводить в течение многих часов и даже суток, если содержать нервно-мышечный препарат в физиологическом растворе или постоянно смачивать его этим раствором, что сохраняет его жизнедеятельность.
Одиночное сокращение и тетанус
В естественных условиях сокращения скелетных мышц вызываются нервными импульсами или волнами возбуждения из центральной нервной системы, поступающими к ним по двигательным нервам. Через мионевральные аппараты волны возбуждения переходят на все мышечные волокна, по которым они распространяются с меньшей скоростью, чем по нервам. В мышечном волокне возбуждение распространяется по особой проводящей системе при участии ионов Са.
При непосредственном приложении раздражителя к мышце, или прямом раздражении, можно вызвать ее сокращение и после перерождения двигательного нерва, после отравления мионевральных аппаратов ядами, прекращающими передачу возбуждения с нервных волокон на мышечные. При непрямом раздражении, когда раздражитель действует на двигательный нерв, достаточен более слабый раздражитель, так как возбудимость нерва больше, чем мышцы. Если мышца подвергается в лабораторных условиях кратковременному одиночному раздражению достаточной интенсивности, то через некоторый промежуток времени после волны возбуждения, называемый скрытым, или латентным, периодом, возникает волна сокращения мышцы. Продолжительность латентного периода больше при непрямом раздражении, чем при прямом, так как затрачивается время, необходимое для проведения возбуждения по нервным волокнам и для прохождения возбуждения через мионевральные аппараты, Имеет значение и способ регистрации этого периода, например, при фотографической регистрации у лягушки он равен 0,0025 с, а при записи миографом — 0,01 с. Продолжительность латентного периода у гладких мышц значительно больше, чем у поперечнополосатых. В скелетных мышцах человека и позвоночных животных этот период длится тысячные доли секунды. Продолжительность латентного периода неодинакова у разных животных. Она зависит и от силы раздражителя.
Чем сильнее раздражитель, тем при равных условиях короче латентный период.
При одиночном сокращении вслед за чрезвычайно коротким латентным периодом наступает короткий период — сокращения. У человека максимум одиночного сокращения мышцы, отводящей большой палец, достигается через 0,05 с.
Скорость распространения волны сокращения у разных животных и у человека неодинакова: у лягушки — около 1м/с, у кролика — 5-10 м/с, у человека — 4-5 м/с. Длительность периода сокращения различна у разных животных и в разных мышцах одного и того же животного. Например, длительность сокращения поперечнополосатой мышцы крыла насекомых — 0,003-0,004 с, мышцы кролика — 0,07 с, лягушки — 0,1 с, черепахи — 4 с. Период сокращения красных мышц кролика больше, чем белых. У лягушки сгибатели сокращаются быстрее разгибателей. Медленно сокращающиеся мышцы способны длительно поддерживать сокращение. У большинства животных с постоянной температурой тела этот период короче (меньше 0,1 с), чем у большинства животных с переменной температурой тела, у которых он может достигать 1 с. Его продолжительность различна у разных видов беспозвоночных. У некоторых насекомых мышцы сокращаются очень часто: у мухи 350 раз в 1 с, у пчелы — 400 раз в 1 с. Длительность сокращения одной и той же мышцы больше в зародышевом периоде и после рождения. С возрастом период одиночного сокращения мышцы постепенно уменьшается. Более быстрые движения свойственны взрослым. Следовательно, продолжительность сокращения скелетных мышц изменяется в фило- и онтогенезе.
Продолжительность сокращения одной и той же мышцы зависит также от ее функционального состояния, от температуры и других условий. При утомлении она увеличивается, а при повышении температуры уменьшается.
Существует зависимость между силой раздражения и величиной (высотой) сокращения скелетной мышцы. При непрямом раздражении нервно-мышечного препарата очень слабым электрическим током вначале мышца не сокращается. Эта сила тока называется подпороговой, или сублиминалъной. Затем по мере увеличения силы тока получается едва заметное сокращение мышцы при минимальной силе тока. Это пороговое, или лиминальное сокращение. При дальнейшем увеличении силы тока высота сокращения мышцы, или степень ее укорочения, все более увеличивается и, наконец, достигает наибольшей величины. Наибольшее сокращение называется максимальным. Все сокращения, лежащие между пороговым и максимальным сокращениями, называются надпороговыми, или субмаксимальными.
После латентного периода и сокращения наступает период расслабления мышцы. Продолжительность этого периода также отличается у разных видов животных, у разных мышц и зависит от функционального состояния мышцы и возраста.
В скелетных мышцах животных с постоянной температурой тела период расслабления продолжается тысячные доли секунды, у животных с переменной температурой тела — сотые и десятые доли секунды, а в гладких мышцах — десятки секунд или даже больше минуты. В естественных условиях одиночные сокращения мышц не происходят. Установлено, что в естественных условиях длительные сокращения мышц образуются вследствие слияния многих одиночных сокращений в результате поступления по двигательному нерву из центральной нервной системы целого ряда или даже больше минуты. В естественных условиях одиночные сокращения мышц не происходит. Установлено, что в естественных условиях длительное сокращение мышц образуются вследствие слияния многих одиночных сокращений в результате поступления по двигательному нерву из центральной нервной системы целого ряда или залпа нервных импульсов. У человека самое большое движение продолжается больше 0,1 с., а непрерывное сокращение мышц рук, ног и туловища может длиться в течении многих минут.
Такое сильное и длительное сокращение скелетной мышцы называется тетаническим сокращением, или тетанусом.
У человека тетаническое сокращение скелетных мышцы вызывается залпом нервных импульсов с частотой не меньше 30, а обычно 50-70 в секунду, при произвольном сокращении, а при напряжении — 5-25. Наиболее благоприятный ритм нервных импульсов, вызывающий у человека максимальное тетаническое сокращение скелетной мышцы, колеблется от 100 до 200 в секунду. У икроножной мышцы лягушки тетаническое сокращение получается при частоте нервных импульсов не ниже 20-30 в секунду. Мышцы же некоторых насекомых дают одиночные сокращения при частоте нервных импульсов до 300 в секунду, и тетанус происходит при более частой нервной импульсации.
Механизм образования тетануса выяснен в эксперименте.
Когда промежутки времени между раздражениями двигательного нерва превышают длительность одиночного сокращения, скелетная мышца успевает сократиться и расслабиться. В этом случае в эксперименте наблюдаются одиночные мышечные сокращения. Когда же эти промежутки времени уменьшаются, то мышца не успевает расслабиться и новые нервные импульсы вызывают суммирование сокращений. Сокращения в этом случае накладываются друг на друга таким образом, что еще различаются отдельные сокращения, вызванные каждым нервным импульсом. Такое накладывание сокращений друг на друга называется суперпозицией, а запись суммированных, но еще различимых сокращений называется неполным, или зубчатым, тетанусом. В этом случае мышца полностью не расслабляется между поступлениями отдельных нервных импульсов.
При дальнейшем учащении нервных импульсов мышца не успевает начать расслабление и происходит полное слияние вершин отдельных сокращений. В этом случае результатом суперпозиции является полный, или гладкий, тетанус — длительное суммированное сокращение. На записи гладкого тетануса невозможно различить отдельные сокращения. Чем меньше длительность одиночного сокращения, тем больше частота импульсов при гладком тетанусе. У кошки длительность одиночного сокращения внутренней прямой мышцы глаза 7,5 мс и гладкий тетанус возникает при 350 импульсах в 1 с; в икроножной мышце — 20 мс и 100 ими/с; в камбаловидной — 100 мс и 30 имп/с.
Г. Гельмгольц считал, что одиночные сокращения при образовании тетануса не влияют друг на друга и последующие сокращения не зависят от предыдущих. Н. Е. Введенский экспериментально доказал, что при образовании тетануса происходит изменение сокращений, из которых он складывается. Каждое сокращение зависит от предыдущего и влияет на последующее. Он обнаружил, что высота тетануса изменяется в зависимости от частоты нервных импульсов и их силы, а также функционального состояния мышцы в момент поступления к ней очередного нервного импульса (1886).
Следовательно, Н. Е. Введенский установил, что тетанус не является простым результатом суперпозиции, как предполагал Г. Гельмгольц.
Изотоническое и изометрическое сокращения
При поступлении в мышцу нервных импульсов по двигательному нерву (непрямое раздражение) или при непосредственном действии на мышцу раздражающего агента (прямое раздражение) в мышце развивается напряжение и она становится короче и толще, объем ее почти не меняется. Это изменение напряжения или формы мышцы называется мышечным сокращением. Если мышца сокращается при отсутствии сопротивления изменению ее длины или при отсутствии груза, то напряжение ее мышечных волокон не изменяется, оно приближается к нулю. Такое сокращение называется изотоническим.
Следует учесть, что и при отсутствии груза естественные сокращения мышц не являются изотоническими, так как при сокращении мышцы поднимают кости и свой собственный вес и, кроме того, преодолевают сопротивление противоположных мышц. К изотоническому приближается сокращение мышц языка, поднимающего только свой вес. Если мышца не может сократиться при непрямом и прямом раздражении вследствие того, что к ней прикреплен чрезмерно большой груз или ее сухожилия прочно закреплены в обоих концах, то в ней развивается максимальное напряжение, а длина ее остается неизменной. Такое сокращение называется изометрическим. Полностью изометрических естественных сокращений также не бывает, так как для измерения напряжения необходимо установить величину максимального груза, сдвигаемого мышцей. Следовательно, мышца при этом сокращается, и ее длина весьма незначительно изменяется. В организме мышцы сокращаются с изменением длины и с напряжением. Такие сокращения называются ауксотоническими (смешанными).
Взаимодействие нервной и гормональной регуляций
Нервная система регулирует поступление в кровь гормонов и обеспечивает единство и взаимодействие всех органов двумя физиологическими механизмами: нервным и нервно-гуморальным, выполняя таким образом ведущую роль в осуществлении единства функций организма в его взаимоотношениях с внешним миром. Нервный и нервно-гуморальный механизмы связаны друг с другом и функционируют одновременно. Нервная система регулирует функции желез внутренней секреции, а гормоны действуют на нервную систему.
МЕТОДИЧЕСКИЕ РАЗРАБОТКИ ДЛЯ ВЫПОЛНЕНИЯ ЛАБОРАТОРНЫХ РАБОТ ПО ФИЗИОЛОГИИ ЖИВОТНЫХ
ПРЕДИСЛОВИЕ
Изучение физиологии животных предполагает усвоение студентами как теоретических знаний, так и овладение навыками экспериментальной работы. Первые приобретаются в ходе лекций и самостоятельной работы с учебным материалом, вторые – на лабораторных занятиях.
Лабораторное занятие предусматривает самостоятельное выполнение студентами экспериментов и интерпретацию получаемых данных. В результате выполнения экспериментов студенты знакомятся с методиками измерения физиологических величин, в том числе и с теми, которые применяются в спортивной и клинической практике. Анализ итоговых фактических материалов позволит студентам приобретать навыки научного мышления, рационального представления и корректной интерпретации данных.
В настоящем учебном пособии приведены подробные описания экспериментов (по всем разделам физиологии человека и животных), которые студенты выполняют во время лабораторных занятий. Постановка опытов описана в форме инструкций, последовательно излагающих этапы их проведения. В конце каждой работы студенту предлагается сделать вывод, что способствует более успешному усвоению материала.
К каждому разделу предлагаются контрольные вопросы, задачи, тестовые задания, которые облегчают самостоятельную подготовку обучающихся и контроль усвоения материала, а также дополнительный блок информации.
Лабораторные опыты адаптированы к программе факультета биологии и биотехнологии по физиологии человека и животных. Включены как классические, так и новые лабораторные опыты.
Основная и дополнительная литература содержит последние издания по всем разделам курса и помогает получить более полные знания по всем вопросам, решаемым в ходе занятий.
Учебное пособие предназначено для студентов специальностей биология, биотехнология, экология. Оно может быть использовано также в системах послевузовского образования и повышения квалификации преподавателей.
РАЗДЕЛ 1. ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Отличительной особенностью высокоспециализированных тканей является способность реагировать на раздражение сложным комплексом физико-химических реакций, называемых возбуждением. Процесс возбуждения связан с наличием в мембране электрически (для Са 2+ и Сl - ) и химически (для Na + и K + ) управляемых каналов, которые могут открываться в ответ на соответствующее раздражение клетки.
Статическая поляризация – наличие разности потенциалов между наружной и внутренней поверхностями клеточной мембраны. В состоянии покоя поверхность клетки всегда электроположительна по отношению к цитоплазме, т.е. поляризована. Эта разность потенциалов, равная
60 мВ, называется потенциалом покоя , или мембранным потенциалом (МП). К факторам, обусловливающим его существование, относятся:
а) наличие К + -Na + трансмембранных градиентов концентрации и градиентов Cl - и Са 2+ ;
б) высокая избирательная проницаемость мембраны для К + , связанная с наличием в ней постоянно открытых в состоянии покоя калиевых каналов. В то же время проницаемость мембраны для Na + в состоянии покоя незначительна. В связи с этим постоянный поток К + из цитоплазмы создает относительный дефицит положительных зарядов на внутренней поверхности и избыток их на наружной поверхности клетки. Токи, регистрируемые в состоянии статической поляризации, называются токами покоя, или калиевыми токами;
Необходимо отметить, что деполяризация может быть как вызванной (при наличии внешнего стимула), так и спонтанной, обусловленной способностью некоторых видов клеток самовозбуждаться. Причиной такого самовозбуждения являются метаболические обменные процессы в самих клетках.
Реполяризация – восстановление исходного уровня МП. Главными факторами, вызывающими реполяризацию мембраны и способствующими восстановлению исходных Na + -К + -градиентов концентрации, являются процессы активного ионного транспорта. Электрографическим проявлением реполяризации является отрицательный следовой потенциал.
Гиперполяризация – увеличение уровня МП. Вслед за восстановлением исходного значения МП (реполяризация) происходит его кратковременное увеличение по сравнению с уровнем покоя, обусловленное повышением проницаемости калиевых каналов и каналов для Cl - . В связи с этим поверхность мембраны приобретает избыточный по сравнению с нормой положительный заряд, а уровень МП становится несколько выше исходного. Электрографическим проявлением гиперполяризации является положительный следовой потенциал. На этом заканчивается одиночный цикл возбуждения.
Изменение возбудимости в различные фазы одиночного цикла возбуждения. Если принять уровень возбудимости в условиях физиологического покоя за норму, то в ходе развития одиночного цикла возбуждения можно наблюдать ее циклические колебания. Так, в период развития начальной деполяризации на очень короткое время возбудимость незначительно повышается по сравнению с исходной. Во время развития полной деполяризации и инверсии заряда возбудимость падает до 0. Время, в течение которого отсутствует возбудимость, называется периодом абсолютной рефрактерности. В это время даже очень сильный раздражитель не может вызвать возбуждение ткани.
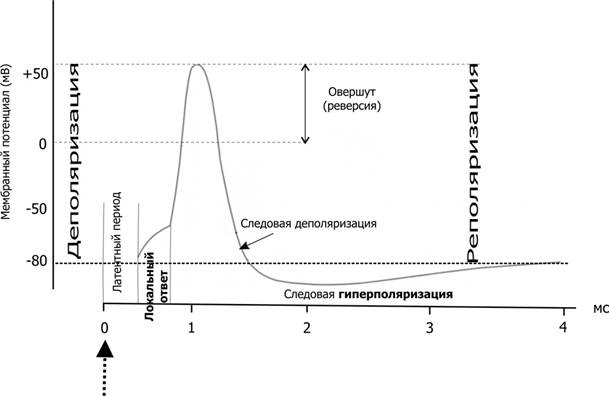
Рис.1 Изменения мембранного потенциала и потенциал действия Вертикальная стрелка в нижней части рисунка — момент появления раздражающего стимула, на отметке –80 мВ — исходный уровень МП.
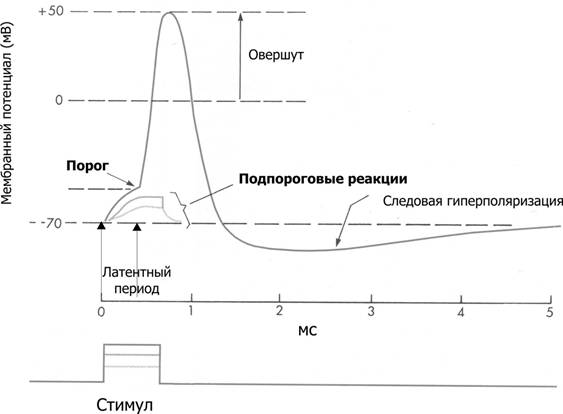
Рис.2 Мембранный потенциал и потенциал действия. На отметке –70 мВ — исходный уровень МП; значение порога показано стрелкой в левой части рисунка, приведены записи подпороговых изменений МП
Изменение возбудимости в различные фазы одиночного цикла возбуждения. Если принять уровень возбудимости в условиях физиологического покоя за норму, то в ходе развития одиночного цикла возбуждения можно наблюдать ее циклические колебания. Так, в период развития начальной деполяризации на очень короткое время возбудимость незначительно повышается по сравнению с исходной. Во время развития полной деполяризации и инверсии заряда возбудимость падает до 0. Время, в течение которого отсутствует возбудимость, называется периодом абсолютной рефрактерности. В это время даже очень сильный раздражитель не может вызвать возбуждение ткани. Фармакологические вещества, увеличивающие время деполяризации и соответственно период абсолютной рефрактерности ткани, вызывают торможение физиологической деятельности. И наоборот, вещества, уменьшающие время деполяризации, оказывают возбуждающее физиологическое действие.
В фазе восстановления МП возбудимость также начинает восстанавливаться, но она еще ниже исходного уровня. Время восстановления ее от 0 до исходной величины называется периодом первичной относительной рефрактерности. Ткань может ответить возбуждением только на сильные, надпороговые, раздражения.
Вслед за периодом относительной рефрактерности наступает короткий период экзальтации – повышенной (по сравнению с исходной) возбудимости. По времени он соответствует процессу реполяризации.
Заключительный этап одиночного цикла возбуждения – повторное снижение возбудимости ниже исходного уровня (но не до 0), называемое периодом вторичной относительной рефрактерности. Он совпадает с развитием гиперполяризации мембраны. Фармакологические вещества, усиливающие гиперполяризацию мембраны, вызывают ослабление физиологической деятельности (гиперполяризационное торможение). Возбуждение может возникнуть только в том случае, если сила раздражения значительно превысит пороговую. После этого возбудимость восстанавливается, и клетка готова к осуществлению следующего цикла возбуждения.
Вопросы для самоподготовки. 1. Критерии оценки функционального состояния ткани. Определение понятий: возбудимость, порог возбуждения, функциональная лабильность. 2. Основные физиологические состояния биологической мембраны. 3. Электрофизиологическая характеристика статической поляризации. Потенциал покоя. 4 Электрофизиологическая характеристика деполяризации. Потенциал действия. 5. Электрофизиологическая характеристика реполяризации и гиперполяризации. Следовые потенциалы.
РАБОТА 1. Приготовление нервно-мышечного препарата лягушки
Нервно-мышечный препарат является универсальным для исследования функций скелетных мышц и нервов, классическим нервно-мышечным препаратом считают икроножную мышцу и седалищный нерв, который ее иннервирует. Работа выполняется на обездвиженной лягушке.
Способы обездвиживания лягушки. Одним из способов обездвиживания лягушки является наркотизация. В качестве наркотического вещества чаще всего используют эфир. Обездвижить лягушку можно также путем введения в подкожный лимфатический мешок миорелаксантов, которые нарушают передачу возбуждения с нерва на мышцу, в связи с чем возникает расслабление скелетных мышц.
Чаще всего обездвиживание лягушки производят путем разрушения спинного и головного мозга. Разрушение ЦНС производят двумя способами – с сохранением головы и путем декапитации. В обоих случаях лягушку завертывают в марлевую салфетку так, чтобы передние лапки оказались прижатыми к туловищу, а задние находились в вытянутом состоянии. При первом способе обездвиживания лягушку держат в левой руке, указательным пальцем которой сгибают голову лягушки так, чтобы между головой и позвоночником образовался некоторый угол. Препаровальной иглой с небольшим нажимом проводят по средней линии головы сверху вниз пока игла не попадет в небольшую ямку – субокципитальное отверстие. Проколов кожу и мембрану в этом месте, иглу вводят на 1-2 мм в полость черепа и несколькими движениями разрушают головной мозг. Слегка извлекая иглу, направляют ее в позвоночный канал и вращательными движениями разрушают спинной мозг. Критерием разрушения является полная релаксация (расслабление) всех групп мышц и отсутствие защитных двигательных рефлексов на пощипывание кожи.
Декапитацию производят введением одной бранши ножниц в ротовую полость и отсечением челюсти и переднего отдела мозга. Разрез должен пройти сразу же за глазными буграми. В открывшийся позвоночный канал вводят препаровальную иглу и разрушают спинной мозг.
Для работы необходимо: препаровальный набор инструментов, лоток, салфетки, операционный столик, раствор Рингера, лягушка
Ход работы. Обездвиженной лягушке перерезать позвоночник с окружающими его тканями на 1-1,5 см выше копчика (рис.3).В руке должны остаться задние лапки с тазовой костью и небольшим отделом позвоночного столба. Захватив край кожи пинцетом, снять кожу с лапок. Удалить внутренности. Получают препарат задних лапок лягушки. Вырезать копчик (уростиль). Для этого, держа препарат так, чтобы лапки висели вниз под прямым углом к позвоночнику, вырезают копчиковую кость, которая при таком положении препарата выдается кверху. Перевернуть препарат на
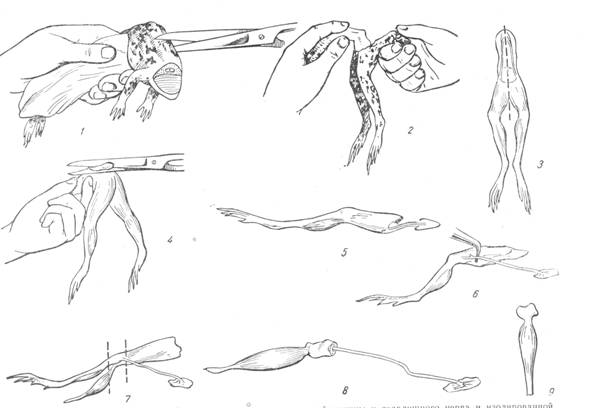
Рис.3. Приготовление нервно-мышечного препарата икроножной мышцв и седалищного
нерва и изолированной икроножной мышцы.
вентральную поверхность и разрезать позвоночник и лонное сращение на две половины (не повредить нервные стволы крестцового сплетения) с тем. Чтобы отделить лапки друг от друга. Получают препараты двух задних лапок.
Следующим этапом является препаровка икроножной мышцы и седалищного нерва. Под ахиллово сухожилие икроножной мышцы подвести лигатуру и перевязать. Затем дистальнее этого места перерезать. Подняв за лигатуру мышцу, отпрепаровывают ее от окружающих тканей. Перерезать берцовую кость ниже коленного сустава, а бедренную – выше его.
Для препаровки седалищного нерва перевернуть препарат дорсальной поверхностью кверху. Стеклянным крючком отодвинуть кверху двуглавую мышцу, а книзу – полуперепончатую. После разрыва фасций между ними обнажается борозда, в которой проходит седалищный нерв. С помощью стеклянных крючков препарируют нерв на всем протяжении до коленного сустава и до позвоночника, оставляя кусочек позвоночника в соединении с нервом. Получают нервно-мышечный препарат.
Следует помнить, что при небрежном приготовлении препарата (прикосновение к нерву руками или металлическим предметом, натяжение, подсыхание, механическое повреждение) он быстро теряет активность Необходимо часто увлажнять препарат раствором Рингера.
Рекомендации к оформлению работы. Зарисовать основные этапы приготовления нервно-мышечного препарата лягушки.
Дата добавления: 2018-02-28 ; просмотров: 1056 ;
Нервно-мышечная система
Методы исследования свойств мышц. Физиологические свойства мышечной ткани обычно изучают на нервно-мышечном препарате лягушки, чаще всего на изолированной икроножной мышце с седалищным нервом. Нервно-мышечный препарат помещают в специальную камеру, в которой поддерживают определенную влажность.
В качестве раздражителя преимущественно используют электрический ток. Однако можно применять механические (щипок, укол, удар), химические (кислоты), температурные (тепло, холод) и другие раздражители.
О функциональном состоянии мышцы судят по результатам регистрации ее Механической реакции (миография) и биопотенциалов (электромиография).
Регистрация мышечного сокращения. Для записи сокращений мышцы ее укрепляют в вертикальном миографе, специальном зажиме, в который помещают бедренную кость нервно-мышечного препарата. Сухожилие мышцы соединяют с пишущим рычажком, свободный конец которого прикладывают к барабану кимографа. Главную часть кимографа - барабан - покрывают бумагой. Барабан приводят в равномерное вращение часовым механизмом или электромотором. Часовой механизм заводят с помощью ключа. Для приведения барабана во вращение смещают стопор, находящийся на стенке коробки часового механизма. Скорость вращения регулируют с помощью специального валика, расположенного на оси барабана. Для замедления вращения применяют крылатки, оказывающие сопротивление действию часового механизма.
Для электрического раздражения мышцы используют электронные стимуляторы или санный аппарат Дюбуа-Реймона.
Основной частью стимулятора является генератор колебаний, включающий электронные лампы или транзисторы. Прибор питается от сети переменного тока и имеет гнезда для подключения электродов. На лицевой панели стимулятора имеются ручки и шкалы. Поворотом тумблера "частота" устанавливают нужную частоту раздражения в секунду, переключателем "длительность" устанавливают необходимую продолжительность стимуляции. С помощью ручки "амплитуда" и тумблера переключения диапазонов амплитуды (×1, ×10) можно изменять величину раздражающего стимула. Имеется тумблер "пуск", при включении которого раздражение подается к препарату. Корпус заземляют.
При раздражении нервно-мышечного препарата возникает мышечное сокращение, которое регистрируется на барабане кимографа в виде кривой, получившей название миограммы.
Для работы необходимы: набор препаровальных инструментов (ножницы большие и малые, пинцет хирургический и глазной, металлический зонд для разрушения спинного мозга, два стеклянных крючка), изоосмотический раствор хлорида натрия для холоднокровных.
Ход работы. 1. Лягушку завернуть в марлевую салфетку, одну браншу ножниц ввести в полость рта, вторую поместить за глазами и, сомкнув их, удалить головной мозг. Зондом разрушить спинной мозг. Острием ножниц проткнуть полость тела, ввести в нее браншу ножниц и, отступя на 1·10 -2 м (1 см) от копчиковой кости, перерезать позвоночник.
2. Держа лягушку за задние лапки и опустив ее головой вниз, отрезать всю свисающую половину туловища и внутренности.
3. Захватить одной рукой остаток позвоночника, другой - край кожи со спины, быстрым движением снять кожу с обеих лапок (в этом случае получают препарат двух задних лапок лягушки).
4. Фиксировать препарат двух лапок так, чтобы они висели вниз под прямым углом к позвоночнику, ножницами осторожно вырезать копчиковую часть. Затем продольно по средней линии отделить лапки друг от друга.
5. Подведя под ахиллово сухожилие браншу ножниц, отделить его по всей длине и, захватив сухожилие пинцетом, оттянуть мышцу в сторону и разорвать фасции, связывающие ее с другими тканями.
6. Для выделения нерва бедро расположить задней поверхностью кверху. Мышцы развести и отпрепарировать двумя стеклянными крючками нерв по всей его длине.
7. Перерезать бедренную кость выше коленного сустава, а кости голени ниже него. Полученный нервно-мышечный препарат поместить в изоосмотический раствор хлорида натрия для холоднокровных.
Для работы необходимы: электронный стимулятор (или санный аппарат Дюбуа-Реймона), электроды, набор препаровальных инструментов, миограф, кимограф, гальванический пинцет, изоосмотический раствор хлорида натрия для холоднокровных.
Ход работы. 1. Приготовить нервно-мышечный препарат лягушки и фиксировать его в миографе.
2. Соединить сухожилие мышцы с пишущим рычажком.
3. Зарядить пишущий рычажок чернилами.
4. Записать мышечные сокращения при раздражении мышцы непосредственно - прямое раздражение и через нерв - непрямое раздражение: а) гальваническим пинцетом; б) электрическим током; в) щипком.
5. Начиная с минимальной (подпороговое раздражение), изменять силу тока и получить минимальное и максимальное сокращение мышцы.
6. Записать сокращение мышцы при минимальной скорости вращения барабана кимографа.
Для работы необходимы: электронный стимулятор (санный аппарат Дюбуа-Реймона), электроды, миограф, кимограф, универсальный штатив, писчики, влажная камера, набор препаровальных инструментов, изоосмотический раствор хлорида натрия для холоднокровных.
Ход работы. 1. Приготовить нервно-мышечный препарат лягушки, укрепить его в миографе, соединить с писчиком. Подвести электроды вплотную к мышце.
2. При частоте раздражения 1 Гц (одиночное раздражение) определить пороговую силу раздражения и несколько увеличить ее.
3. Записать на барабане кимографа миограммы при нанесении на мышцу раздражения: а) 1 Гц; б) 5-10 Гц; в) 20-40 Гц.
Для работы необходимы: электронный стимулятор (катушка Дюбуа-Реймона), влажная камера с электродами, миограф с подпоркой, набор грузов с крючками, набор препаровальных инструментов, изоосмотический раствор хлорида натрия для холоднокровных.
Ход работы. 1. Приготовить нервно-мышечный препарат лягушки.
2. Изолировать икроножную мышцу.
3. Ахиллово сухожилие соединить с рычагом миографа прочной ниткой или проволокой. Под рычаг поставить особую подпорку, которая будет препятствовать растягиванию мышцы при подвешивании груза и позволит лишь поднимать груз при сокращении.
4. Подобрать силу и частоту раздражений, дающих наиболее высокое тетаническое сокращение. Запись вести на неподвижном барабане кимографа.
5. Записать максимальную высоту сокращения мышцы, без нагрузки.
6. К рычагу (к той же точке, за которую тянет мышца) подвесить груз в 2,5·10 -2 -3·10 -2 кг (25-30 г) и снова записать высоту сокращения в ответ на раздражение. Под графиком отметить массу груза.
7. Не меняя силу и частоту раздражения, при постепенном увеличении нагрузки на 5·10 -3 кг (5 г) и одновременной регистрации высоты сокращения найти предельный груз, который мышца может поднять. Эта величина, выраженная в килограммах, и будет соответствовать абсолютной силе мышцы.
8. Вычислить работу мышцы при различных нагрузках по формуле:
где А - работа; m - масса груза; h - высота сокращения.
Для работы необходимы: эргограф, кимограф, метроном, набор грузов (1-3 кг), циркуль, линейка.
Ход работы. 1. Укрепить предплечье в станке эргографа.
2. Надеть кожаное кольцо, соединенное с грузом и пишущим устройством, на указательный палец. Остальными пальцами для исключения их движения охватить вертикальный стержень.
3. Подвесить на шнур груз массой 2 кг и включить метроном с частотой 60 ударов в 60 с (1 мин).
4. Обследуемый начинает поднимать груз в заданном ритме до полного утомления.
5. Отметить время начала и окончания работы и определить ее продолжительность.
6. По эргограмме определить величину работы, выполненной сгибателем пальца. Для этого определить высоту каждого подъема кривой, полученные величины сложить и умножить на массу груза.
7. После отдыха повторить исследование с грузом массой 3 кг и частотой совершаемых движений 120 в 60 с (1 мин).
Для работы необходим: кистевой динамометр.
Ход работы. 1. Определить мышечную силу сгибателей пальцев правой и левой руки с помощью кистевого динамометра.
2. Определить силу обеих рук на растяжение.
3. Определить силу разгибателей пальцев после многократных сжатий пружины динамометра.
Читайте также:
