
Механизм передачи нервного импульса физиология
·химический — это место близкого прилегания двух нервных клеток, для передачи нервного импульса через которое клетка-источник выпускает в межклеточное пространство особое вещество, нейромедиатор, присутствие которого в синаптической щели возбуждает или затормаживает клетку-приёмник.
·электрический (эфапс) — место более близкого прилегания пары клеток, где их мембраны соединяются с помощью особых белковых образований — коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе — 3,5 нм (обычное межклеточное — 20 нм). Так как сопротивление внеклеточной жидкости мало (в данном случае), импульсы через синапс проходят не задерживаясь. Электрические синапсы обычно бывают возбуждающими.
·смешанные синапсы — пресинаптический потенциал действия создает ток, который деполяризуетпостсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Наиболее распространены химические синапсы. Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
По местоположению и принадлежности структурам
·аксо-дендритические — с дендритами, в том числе
·аксо-шипиковые — с дендритными шипиками, выростами на дендритах;
·аксо-соматические — с телами нейронов;
·аксо-аксональные — между аксонами;
·дендро-дендритические — между дендритами;
По нейромедиатору
·аминергические, содержащие биогенные амины (например, серотонин, дофамин);
·в том числе адренергические, содержащие адреналин или норадреналин;
·холинергические, содержащие ацетилхолин;
·пуринергические, содержащие пурины;
·пептидергические, содержащие пептиды.
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По знаку действия
·возбуждающие
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор — глицин) и ГАМК-ергические синапсы (медиатор — гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов: 1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала; 2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение.
В некоторых синапсах присутствует постсинаптическое уплотнение — электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические — симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
№11 Современные представления о медиаторных механизмах, возбуждающих синапсов. Ионный механизм ВПСП
ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
Передача возбуждения в синапсе – сложный физиологический процесс, имеющий несколько стадий:
Синтез медиатора.
Химические медиаторы являются веществами, синтезируемыми в цитоплазме нейронов и нервных окончаниях из компонентов внутри- и внеклеточной жидкостей. Медиаторы обеспечивают одностороннюю передачу возбуждения от нервного волокна к клетке или другому нейрону.
1) В синапсах скелетных мышц вырабатывается только один вид медиатора – ацетилхолин, из холина (продукта печеночной секреции) и уксусной кислоты при участии ферментов, регулирующих активность метаболизма. Норадреналин, как и адреналин, относится к катехоламинам, но вырабатывается и в адренергических синапсах, которые относятся, в основном, к центральным синапсам и нейроорганным синапсам симпатической нервной системы.
2) В синапсах гладких мышц вырабатывается как АХ, так и НА, и эти медиаторы могут оказывать через синапс разное воздействие на гладкую мышцу в зависимости от вида активируемых ими ионных каналов на постсинаптической мембране.
3) В синапсах головного мозга функции медиаторов выполняют более 300 биологически активных веществ. Медиаторы синтезируются постоянно и депонируются в синаптических пузырьках нервных окончаний.
Секреция медиатора.
Дата добавления: 2018-06-01 ; просмотров: 558 ;
Раздражения рецепторов вызывают преобразование воздействующей энергии раздражителя в нервные импульсы, передача которых в нервной системе осуществляется с помощью синапсов.
Функциональные структуры клеточной мембраны. Клеточная мембрана (оболочка клетки) представляет собой тонкую липопротеиновую пластинку, содержание липидов – около 40%, белков – 60%. Схематично клеточную мембрану можно представить следующим образом: мембрана состоит из двойного слоя молекул фосфолипидов, покрытого изнутри слоем белковых молекул, а снаружи – слоем молекул сложных углеводов. В клеточной мембране имеются тончайшие канальцы – ионные каналы, обладающие селективностью. Различают каналы, которые пропускают только один ион (натрия, калия, кальция, хлора), либо несколько.
Потенциал покоя и потенциал действия. В состоянии покоя в протоплазме нервной клетки концентрация ионов калия более, чем в 30 раз превышает концентрацию этих ионов в наружном растворе. Мембрана практически непроницаема для натрия, тогда как калий проходит сквозь нее. Диффузия ионов калия из протоплазмы во внешнюю жидкость очень высокая, что и придает наружной мембране положительный заряд, а внутренней – отрицательный. Таким образом, концентрация ионов калия является основным фактором, формирующим и определяющим величину потенциала покоя (ПП).
При действии на клетку раздражения проницаемость мембраны для ионов натрия резко повышается и становится, примерно, в 10 раз больше проницаемости для ионов калия. Поэтому поток положительно заряженных ионов калия из протоплазмы во внешний раствор уменьшается, а возрастает поток положительно заряженных ионов натрия из внешнего раствора в протоплазму клетки. Это приводит к перезарядке мембраны, наружная поверхность становится заряженной электро-отрицательно, а внутренняя - положительно (фаза деполяризации).
Повышение проницаемости мембраны для ионов натрия продолжается очень короткое время. Вслед за этим в клетке возникают восстановительные процессы, приводящие к тому, что проницаемость для ионов натрия снова понижается, а проницаемость ее для ионов калия возрастает. И в итоге этих двух процессов снова наружная мембрана приобретает положительный заряд, а внутренняя – отрицательный (фаза реполяризации).
Мгновенное увеличение проницаемости для ионов натрия и их проникновение в клетку достаточно, чтобы изменить знак мембранного потенциала и возникает потенциал действия (ПД), который распространяется по аксону с довольно большой скоростью, длительность ПД обычно составляет 1-3 мс.
Синаптическая передача информации. Место передачи возбуждения с одного нейрона на другой называется синапсом (в переводе с греческого – контакт). Синапс представляет собой мембраны двух соседних нейронов (пресинаптическая и постсинаптическая мембраны) и пространство между ними, которое называется синаптической щелью.
Различают аксо-соматические синапсы, сформированные мембранами аксона и телом (сомой) другого нейрона, аксо-дендритные, состоящие из мембраны аксона и дендритами другого нейрона, аксо-аксональные, при которых аксон подходит к аксону другого нейрона. Синапс между аксонами и мышечными волокнами называется нейромышечной пластинкой.
Нервный импульс по аксону достигает окончания аксона и вызывает открытие каналов для кальция на пресинаптической мембране. Здесь, на пресинаптической мембране находятся везикулы (пузырьки), которые содержат биологически активные вещества – медиаторы.
Открытие кальцевых каналов приводит к деполяризации на пресинаптической мембране. Кальций входит в связь с белками, образующими оболочку пузырьков, в которых хранится медиатор. Затем пузырьки лопаются и все содержимое поступает в синаптическую щель. Далее молекулы медиатора связываются со специальными белковыми молекулами (рецепторами), которые находятся на мембране другого нейрона – на постсинаптической мембране.
Когда молекулы медиаторов связываются с рецепторами, то на постсинаптической мембране открываются каналы для ионов натрия и калия, вызывая на ней изменение потенциала (деполяризацию). Этот потенциал получил название – постсинаптический потенциал (ПСП). В зависимости от характера открытых ионных каналов возникает возбудительный (ВПСП) или тормозный (ТПСП) постсинаптические потенциалы
Таким образом, возбуждение (ПД) нейрона в синапсе превращается из электрического импульса в химический импульс (выброс медиатора из везикул).
Время между началом пресинаптической деполяризации и постсинаптической реакцией составляет 0,5 мс, это – синаптическая задержка.
Основные медиаторы: ацетилхолин, моноамины (серотонин, гистамин), катехоламины (дофамин, норадреналин, адреналин), аминокислоты (глутамат, глицин, аспартат, гамма-аминомаслянная кислота – ГАМК, аланин), пептиды, вазопрессин, окситоцин, аденозин, АТФ и др.
Спинной мозг

Спинной мозг, по внешнему виду представляет собой длинный, цилиндрической формы, уплощенный спереди назад тяж. В связи с этим поперечный диаметр спинного мозга больше переднезаднего.
Спинной мозг располагается в позвоночном канале и на уровне нижнего края большого затылочного отверстия переходит в головной мозг. В этом месте из спинного мозга (верхняя его граница) выходят корешки, образующие правый и левый спинномозговые нервы. Нижняя граница спинного мозга соответствует уровню 1-11 поясничных позвонков. Ниже этого уровня верхушка мозгового конуса спинного мозга продолжается в тонкую терминальную (концевую) нить. Терминальная нить в своих верхних отделах еще содержит нервную ткань и представляет собой рудимент каудального конца спинного мозга. Эта часть терминальной нити, получившая название внутренней, окружена корешками поясничных и крестцовых спинномозговых нервов и вместе с ними находится в слепо заканчивающемся мешке, образованном твердой оболочкой спинного мозга. У взрослого человека внутренняя часть терминальной нити имеет длину около 15 см. Ниже уровня 2 крестцового позвонка терминальная нить представляет собой соединительнотканное образование, являющееся продолжением всех трех оболочек спинного мозга и получившее название наружной части терминальной нити. Длина этой части около 8 см. Заканчивается она на уровне тела 2 копчикового позвонка, срастаясь с его надкостницей.
Длина спинного мозга у взрослого человека в среднем 43 см (у мужчин 45 см, у женщин 41-42 см), масса - около 34-38 г, что составляет примерно 2 % от массы головного мозга.
В шейном и поясничнокрестцовом отделах спинного мозга обнаруживаются два заметных утолщения: шейное утолщение и пояснично-крестцовое утолщение. Образование утолщений объясняется тем, что от шейного и поясничнорестцового отделов спинного мозга осуществляется иннервация соответственно верхних и нижних конечностей. В этих отделах в спинном мозге имеется большее по сравнению с другими отделами количество нервных клеток и волокон. На нижних отделах спинной мозг постепенно суживается и образует мозговой конус.
На передней поверхности спинного мозга видна передняя срединная щель, которая вдается в ткань спинного мозга глубже, чем задняя срединная борода. Эти борозды являются границами, разделяющими спинной мозг на две симметричные половины. В глубине задней срединной борозды имеется проникающая почти во всю толщу белого вещества глиальная задняя срединная перегородка. Эта перегородка доходит до задней поверхности серого вещества спинного мозга.
На передней поверхности спинного мозга, с каждой стороны от передней щели, проходит переднелатеральная борода. Она является местом выхода из спинного мозга передних (двигательных) корешков спинномозговых нервов и границей на поверхности спинного мозга между передним и боковым канатиками. На задней поверхности на каждой половине спинного мозга имеется заднелатеральная борозда, место проникновения в спинной мозг задних чувствительных корешков спинномозговых нервов. Эта борозда служит границей между боковым и задним канатиками.
Передний корелток состоит ив отростков двигательных (моторных) нервных клеток, расположенных в переднем роге серого вещества спинного мозга. Задний корешок - чувствительный, представлен совокупностью проникающих в спинной мозг центральных отростков псевдоуниполярных клеток, тела которых образуют спинномозговой узел, лежащий у места соединения заднего корешка с передним. На всем протяжении спинного мозга с каждой его стороны отходит 31 пара корешков. Передний и задний корешки у внутреннего края межпозвоночного отверстия сближаются, сливаются друг с другом и образуют спинномозговой нерв. Таким образом, ив корешков образуется 31 пара спинномозговых нервов. Участок спинного мозга, соответствующий двум парам корешков (два передних и два задних), называют сегментон.
Для врача очень важно знать топографические взаимоотношения сегментов спинного мозга с позвоночным столбом (скелетотопия сегментов) . Протяженность спинного мозга значительно меньше длины позвоночного столба, поэтому порядковый номер какого-либо сегмента спинного мозга и уровень его положения, начиная с нижнего шейного отдела, не соответствует порядковому номеру одноименного позвонка. Положение сегментов по отношению к позвонкам можно определить следующим образом. Верхние шейные сегменты расположены на уровне соответствующих их порядковому номеру тел позвонков. Нижние шейные и верхние грудные сегменты лежат на один по- звонок выше, чем тела соответствующих позвонков. В среднем грудном отделе эта разница между соответствующим сегментом спинного мозга и телом позвонка увеличивается уже на 2 позвонка, в нижнем грудном - на З. Поясничные сегменты спинного мозга лежат в позвоночном канале на уровне тел 10, 11 грудных позвонков, крестцовые и копчиковый сегменты - на уровне 12 грудного и 1 поясничного позвонков.
Спинной мозг состоит из нервных клеток и волокон серого вещества, имеющего на поперечном срезе вид буквы В или бабочки с расправленными крыльями. За периферии от серого вещества находится белое вещество, образованное только нервными волокнами.
В сером веществе спинного мозга имеется центральный канал. Он является остатком полости нервной трубки и содержит спинномозговую жидкость. Верхний конец канала сообщается с 9 желудочком, а нижний, несколько расширяясь, образует слепо заканчивающийся концевой желудочек. Стенки центрального канала спинного мозга выстланы эпендимой, вокруг которой находится центральное студенистое (серое) вещество. У взрослого человека центральный канал в различных отделах спинного мозга, а иногда и на всем протяжении зарастает.
Серое вещество, на протяжении спинного мозга справа и слева от центрального канала образует симметричные серые столбы. Кпереди и кзади от центрального канала спинного мозга эти серые столбы связаны друг с другом тонкими пластинками серого вещества, получившими название передней и задней спаек.
В каждом столбе серого вещества различают переднюю его часть - передний столб и заднюю часть - задний столб. За уровне нижнего шейного, всех грудных и двух верхних поясничных сегментов спинного мозга.
Серое вещество с каждой стороны образует боковое выпячивание - боковой столб. В других отделах спинного мозга (выше 8 шейного и ниже 2 поясничного сегментов) боковые столбы отсутствуют.
За поперечном срезе спинного мозга столбы серого вещества с каждой стороны имеют вид рогов. Выделяют более широкий передний рог и узкий задний рог1, соответствующие переднему и заднему столбам. Боковой рог, соответствует боковому промежуточному столбу (автономному) серого вещества.
В передних рогах расположены крупные нервные корешковые клетки - двигательные (эфферентные) нейроны. Эти нейроны образуют 5 ядер: два латеральных (передне и заднелатеральное), два медиальных (передне и заднемедиальное) и центральное ядро. Задние рога спинного мозга представлены преимущественно более мелкими клетками. В составе задних, или чувствительных, корешков находятся центральные отростки псевдоуниполярных клеток, расположенных в спинномозговых (чувствительных) узлах.
Серое вещество задних рогов спинного мозга неоднородно. Основная масса нервных клеток заднего рога образует собственное его ядро. В белом веществе, непосредственно примыкающем к верхушке заднего рога серого вещества, выделяют пограничную зону. Кпереди от последней в сером веществе расположена губчатая зона, которая получила свое название в связи с наличием в этом отделе крупнопетлистой глиальной сети, содержащей нервные клетки. Еще более кпереди выделяется студенистое вещество, состоящее из мелких нервных клеток. Отростки нервных клеток студенистого вещества, губчатой зоны и диффузно рассеянных во всем сером веществе пучковых клеток осуществляют связь с несколькими соседними сегментами. Как правило, они заканчиваются синапсами с нейронами, расположенными в передних рогах своего сегмента, а также выше и нижележащих сегментов. Направляясь от задних рогов серого вещества к передним рогам, отростки этих клеток располагаются по периферии серого вещества, образуя возле него узкую каемку белого вещества. Эти пучки нервных волокон получили название передних, латеральных и задних собственных пучков. Клетки всех ядер задних рогов серого вещества - это, как правило, вставочные (промежуточные, или кондукторные) нейроны. Нейриты, отходящие от нервных клеток, совокупность которых составляет центральное и грудное ядра задних рогов, направляются в белом веществе спинного мозга к головному мозгу.
Промежуточная зона серого вещества спинного мозга расположена между передним и задним рогами. Здесь на протяжении с 8 шейного по 2 поясничный сегмент имеется выступ серого вещества - боковой рог.
В медиальной части основания бокового рога заметно хорошо очерченное прослойкой белого вещества трудное ядро, состоящее из крупных нервных клеток. Это ядро тянется вдоль всего заднего столба серого вещества в виде клеточного тяжа (ядро Кларка) . Наибольший диаметр этого ядра на уровне от 11 грудного до 1 поясничного сегмента. В боковых рогах находятся центры симпатической части вегетативной нервной системы в виде нескольких групп мелких нервных клеток, объединенных в латеральное промежуточное (серое) вещество. Аксоны этих клеток проходят через передний рог и выходят из спинного мозга в составе передних корешков.
В промежуточной зоне расположено центральное промежуточное (серое) вещество, отростки клеток которого участвуют в образовании спиномозжечкового пути. На уровне шейных сегментов спинного мозга между передним и задним рогами, а на уровне верхнегрудных сегментов - между боковыми и задним рогами в белом веществе, примыкающем к серому, расположена ретикулярная формация. Ретикулярная формация имеет здесь вид тонких перекладин серого вещества, пересекающихся в различных направлениях, и состоит из нервных клеток с большим количеством отростков.
Серое вещество спинного мозга с задними и передними корешками спинномозговых нервов и собственными пучками белого вещества, окаймляющими серое вещество, образует собственный, или сегментарный, аппарат спинного мозга. Основное назначение сегментарного аппарата как филогенетически наиболее старой части спинного мозга - осуществление врожденных реакций (рефлексов) в ответ на раздражение (внутреннее или внешнее). И. П. Павлов определил этот вид деятельности сегментарного аппарата спинного мозга термином "безусловные рефлексы".
Белое вещество, как отмечалось, локализуется кнаружи от серого вещества. Борозды спинного мозга разделяют белое вещество на симметрично расположенные справа и слева три канатика. Передний канатик, находится между передней срединной щелью и передней латеральной бороздой. Н белом веществе кзади от передней срединной щели различают переднюю белую спайку, которая соединяет передние канатики правой и левой сторон. Задний канатик, находится между задней срединной и задней латеральной бороздами. Боковой канатик- это участок белого вещества между передней и задней латеральными бороздами.
1. Механизм проведения возбуждения.
2. Синаптическая передача нервного импульса.
3. Свойства нервных центров.
4. Координирующая роль ЦНС.
5. Торможение в ЦНС.
Механизм проведения возбуждения. Нервные импульсы распространяются по нервным волокнам от рецепторов к нервному центру или от нервного центра к эффектору. Нервные волокна делят на три класса: А, В и С. К классу А относятся толстые миелиновые волокна, скорость проведения возбуждения в них достигает 70-120 м/c. К классу В относятся миелинизированные преганглионарные волокна вегетативной нервной системы, скорость проведения возбуждения в них 3-14 м/c. Волокна класса С – безмиелиновые постганглионарные волокна вегетативной нервной системы. Скорость проведения возбуждения – 0,5-2 м/с.
Механизм проведения возбуждения в нервных волокнах объясняется возникновением локальных токов, появляющихся между возбужденным и невозбужденным участками мембраны нервного волокна.
В безмиелиновых волокнах возбуждение распространяется непрерывно, а в миелинизированных волокнах – скачками между перехватами Ранвье.
Синаптическая передача нервного импульса.Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку). В нервной системе синапсы образуются между отростками разных нейронов, а так же между отростками и телами клеток. Соответственно их называют аксо-аксональными, аксо-дендритными, аксо-соматическими, дендро-дендритными. Передача возбуждения с отростка одной нервной клетки на отросток или тело другой нервной клетки возможна двумя способами: электрическим и химическим. Возбуждающим электрическим синапсам свойственна очень узкая синаптическая щель и очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран для прохождения через них электрического тока. Такие синапсы находятся в сердечной мышце, гладких мышцах и железистой ткани. Основным способом передачи возбуждения между нервными клетками является химический. Синапс с химической передачей состоит из синаптической терминали, пресинаптической мембраны, синаптической щели и постсинаптической мембраны.
В пресинаптическом окончании синапса находятся митохондрии и пресинаптические пузырьки (визикулы) c медиатором. Синаптическая щель имеет ширину 20-50 нм. Постсинаптическая мембрана утолщена и образована мембраной иннервируемой клетки, а в концевой пластинке – мембраной мышечного волокна.
Механизм синаптической передачи.Под влиянием нервного импульса наступает деполяризация окончаний аксона, что повышает в нем концентрацию Ca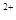
, и содержание синаптических пузырьков выбрасывается в синаптическую щель. Медиатор (ацетилхолин) диффундирует в синаптическую щель и связывается с рецепторами постсинаптической мембраны. Действие молекул медиатора ведет к открытию ионных каналов для Na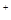
и К
, что вызывает возникновение возбуждающего постсинаптического потенциала (ВПСП). В тормозных синапсах, тормозный медиатор (гамма-аминомасляная кислота) открывает в постсинаптической мембране селективные хлорные каналы. При этом возникает тормозный постсинаптический потенциал (ТПСП) .
Свойства нервных центров.Группа нейронов, необходимых для осуществления определенного рефлекса или управления одной из функций организма, образует нервный центр. Свойства нервных центров зависят от строения и механизма передачи возбуждения в синапсах: 1.Одностороннее проведение возбуждения. В нервном центре, в отличии от нервного волокна, возбуждение распространяется только в одном направлении: от центростремительного нейрона к центробежному. 2. Замедление проведение возбуждения. Это свойство вызвано тем, что в нервном центре может быть несколько нейронов и, следовательно, столько же синапсов. В каждом синапсе происходит синаптическая задержка проведения возбуждения. 3. Суммация. Заключается в накоплении (сложении) эффектов подпороговых раздражений. Одно пдпороговое раздражение не вызывает ответной рефлекторной реакции, а несколько подпороговых раздражений в сумме дают нужный эффект. 4. Трансформация ритма возбуждения. Нервные центры обладают способностью трансформировать частоту и ритм поступающих импульсов. На одиночное раздражение, поступившее в нервный центр, он может ответить серией импульсов. 5. Последействие. Ответная рефлекторная реакция продолжается некоторое время после прекращения действия раздражителя. Причиной последействия являются следовая деполяризация и циркуляция нервных импульсов по кольцевым связям между нейронами данного центра. 6. Утомляемость нервного центра. Утомление в ЦНС возникает в нервном центре. Это обусловлено его низкой лабильностью. Утомление в нервном центре проявляется в постепенном уменьшении, а затем и прекращении рефлекторного ответа при действии раздражителя. Причиной возникновения утомления является нарушение передачи возбуждения в синапсах. 7. Пластичностьнервных центров. В определенных условиях нервные центры перестраиваются и приобретают новые, несвойственные им ранее функции. Это особенно ярко проявляется при операциях удаления различных отделов мозга.
Координирующая роль ЦНС. Приспособление организма к различным изменениям внешней среды возможно благодаря наличию в ЦНС координации функций. Под координацией функций понимают взаимодействие нейронов, а следовательно и нервных процессовв ЦНС, которое обеспечивает ее согласованную деятельность, направленную на интеграцию (объединение) функций различных органов и систем организма. Известен ряд механизмов, лежащих в основе координирующей деятельности нервной системы. Одни из них связаны с морфологическими особенностями ее строения (принцип общего конечного пути, принцип обратных связей), другие – с функциональными свойствами (иррадиация, индукция, доминанта).
Принцип общего конечного пути. Был открыт Ч.С. Шеррингтоном. Этот принцип исходит из анатомического соотношения между афферентными и эфферентными нейронами. Количество чувствительных нейронов, приносящих возбуждение в ЦНС в 5 раз больше, чем двигательных. Поэтому, к одному мотонейрону поступают импульсы от многих рецепторов, расположенных в различных частях тела. Этот процесс называется конвергенцией. Таким образом, самые разнообразные стимулы могут быть причиной одной и той же рефлекторной реакции, т.е. происходит борьба за “общий конечный путь”.
Принцип обратных связей. Воздействие работающего органа на состояние управляющего им центра называется обратной связью. Различают положительные и отрицательные обратные связи. Если импульсы, возникающие в результате какой-либо рефлекторной реакции, поступая в управляющий ею нервный центр, усиливают ее, - это положительная обратная связь. Если они угнетают эту реакцию, то это отрицательная обратная связь.
Иррадиация. Возбуждение, возникшее в одном из нервных центров, способно распространятся по ЦНС, возбуждая новые участки. Процесс распространения возбуждения называют иррадиацией. Она обусловлена наличием многочисленных связей между нейронами в ЦНС.
Индукция. Процессы возбуждения и торможения в ЦНС находятся в определенных отношениях, которые осуществляются по законам индукции. Возбуждение, возникшее в одном нервном центре, “наводит” торможение на другой, и наоборот.
Принципдоминанты. Был разработан А.А. Ухтомским. Господствующий очаг возбуждения, определяющий характер ответных реакций организма на внешние и внутренние раздражения, Ухтомский назвал доминантой. Доминантный очаг возбуждения характеризуется признаками: 1) повышенной возбудимостью; 2) стойкостью возбуждения; 3) повышенной способностью к суммации возбуждения; 4) инерцией, т.е. способностью длительно сохранять возбуждение после окончания действия раздражителя; 5) способностью вызывать сопряженное торможение.
Торможение в ЦНС. Торможение – это особый нервный процесс проявляющийся в уменьшении или полном исчезновении ответной реакции. Процесс торможения в ЦНС был открыт И.М. Сеченовым (1862). В ЦНС наряду с возбуждающими имеются и тормозные нейроны. На каждой нервной клетке располагаются возбуждающие и тормозные синапсы. Поэтому, в каждый момент на теле нейрона в одних синапсах возникает возбуждение, а в других – торможение.
По характеру возникновения различают первичное и вторичное торможение.
Первичное торможение возникает под влиянием раздражения сразу без предварительного возбуждения и осуществляется с участием тормозных синапсов. Вторичное торможение осуществляется без участия тормозных структур и возникает вследствие перехода возбуждения в торможение. Первичное торможение по месту возникновения может быть пресинаптическим и постсинаптическим. Пресинаптическое торможение осуществляется перед прохождением импульса через синапс. Постсинаптическое торможение может быть прямым и возвратным. Прямое торможение осуществляется вставочными нейронами спинного мозга, корзинчатыми нейронами таламуса, тормозными клетками мозжечка, а возвратное торможение через специальные тормозные нейроны – клетки Реншоу.
Читайте также:
