
Особенности проведения нервных импульсов
Проведение нервных импульсов осуществляется при помощи отростков нервных клеток – нервных волокон. По особенностям строения и расположения осевых цилиндров в составе нерва (по отношению к шванновским клеткам) различают волокна мякотного (миелинизированные) и безмякотного (немиелинизированного) типа. Как правило, нервы включают волокна обоих типов в разном количественном соотношении. Миелиновые волокна преобладают в чувствительных и двигательных нервах органов чувств, скелетных мышц и вегетативной нервной системы. Безмиелиновые преимущественно встречаются в составе нервов симпатического отдела.
Волокна мякотного и безмякотного типов состоят из одинаковых структурных компонентов: отростка нервной клетки (осевого цилиндра), шванновских клеток (леммоцитов) и базальной мембраны (тонкой пластинки, состоящей из рыхлой волокнистой соединительной ткани и покрывающей каждое нервное волокно снаружи). Процесс миелинизации (формирование оболочки) продолжается до возраста 25лет, и обеспечивает зрелость НС. Миелиновое волокно представлено осевым цилиндром, окруженным миелиновой оболочкой. Шванновские клетки, окружающие отросток нейроцита, образуют дупликатуры цитоплазма-тической мембраны, называемые мезаксонами. Многократно обертываясь вокруг осевого цилиндра, мезаксон формирует миелиновую оболочку нервного волокна, представляющую собой плотный липидно-белковый футляр. На протяжении мякотного нервного волокна через примерно равные интервалы имеются участки истончения миелинового слоя, ширина которых составляет около 1 мкм. Эти участки получили название перехватов Ранвье. На эти хучастках плотность расположения натриевых ионных каналов в 200 раз больше, чем на соседних участках, поэтому они являются высокочувствительными. Они имеют большое функциональное значение при проведении нервного импульса. Расстояние от одного до другого перехвата называется межузловым сегментом. Мякотные волокна могут располагаться как в пределах центральной нервной системы, так и вне ее. В зависимости от этого среди них различают центральные и периферические. Миелиновые волокна характеризуются высокой скоростью проведения импульсов (60-120м/с). Эта особенность обусловлена изолирующей функцией миелинового слоя и наличием перехватов Ранвье. Нервный импульс в виде ПД в мякотном волокне распространяется скачкообразным способом (сальтоторно). Поэтому распространение нервного импульса по такому волокну проходит быстрее и требует меньшей затраты энергии. Основные функции миелиновой оболочки:
1) изолирующая: оболочка защищает электрические сигналы от перемещения с волокна на волокно (изолированные провода);
2) трофическая – доставка питательных веществ.
Безмякотные нервные волокна построены более примитивно. Они представлены цепочкой шванновских клеток, в которую вдавлено от одного и более осевых цилиндров. В среднем их количество составляет от 5 до 20. Миелинового слоя в этих клетках нет, леммоциты образуют между осевыми цилиндрами мезаксо-ны, не играющие существенной роли в проведении нервного возбуждения. Отростки нейроцитов расположены довольно рыхло и плохо изолированы. Поэтому они могут переходить из одного безмиелинового волокна в другое. Снаружи волокно покрыто соединительно-тканной базальной пластинкой. В связи с перечисленными особенностями строения безмякотные нервные волокна называют волокнами кабельного типа. Скорость проведения в них (3–5 м/с) гораздо ниже, чем в миелиновых. В безмякотном волокне оболочка аксона находится в цитоплазме глиальной клетки. Способ продвижения нервного импульса по такому волокну называется непрерывным. Способ медленный и энергозатратный. Такие волокна в структуре ВНС – сетчатка глаза, гладкие мышцы внутренних органов.
- ЖАНРЫ 360
- АВТОРЫ 262 652
- КНИГИ 606 640
- СЕРИИ 22 752
- ПОЛЬЗОВАТЕЛИ 571 313
Алексей Солодков, Елена Сологуб
Физиология человека. Общая. Спортивная. Возрастная
Учебник для высших учебных заведений физической культуры. 7-е издание
Допущен Министерством РФ по физической культуре и спорту в качестве учебника для высших учебных заведений физической культуры
Издание подготовлено на кафедре физиологии Национального государственного университета физической культуры, спорта и здоровья им. П. Ф. Лесгафта, Санкт-Петербург
В. И. Кулешов, доктор мед. наук, проф. (ВмедА им. С. М. Кирова)
И. М. Козлов, доктор биол. и доктор пед. наук, проф. (НГУ им. П. Ф. Лесгафта, Санкт-Петербург)

Солодков Алексей Сергеевич – профессор кафедры физиологии Национального государственного университета физической культуры, спорта и здоровья им. П. Ф. Лесгафта (в течение 25 лет заведующий кафедрой 1986–2012 гг.).
Доктор медицинских наук, профессор, автор более 490 печатных работ по физиологии и психофизиологии труда, военного труда и спорта, соавтор 13 учебников, 22 учебных и учебно-методических пособий по различным разделам физиологии человека.

Сологуб Елена Борисовна – доктор биологических наук, профессор. С 2002 г. проживает в Нью-Йорке (США).
На кафедре физиологии Национального государственного университета физической культуры, спорта и здоровья им. П. Ф. Лесгафта работала с 1956 г., с 1986 г. по 2002 г. – в должности профессора кафедры. Была избрана академиком Российской академии Медико-технических наук, Почетным работником высшего образования России, членом Правления СПб общества физиологов, биохимиков и фармакологов им. И. М. Сеченова.
Автор около 300 печатных работ по электроэнцефалографии, общей и спортивной физиологии отдельных видов спорта, изданных на русском и иностранных языках.
Физиология человека является теоретической основой целого ряда практических дисциплин (медицины, психологии, педагогики, биомеханики, биохимии и др.). Без понимания нормального течения физиологических процессов и характеризующих их констант различные специалисты не могут правильно оценивать функциональное состояние организма человека и его работоспособность в различных условиях деятельности. Знание физиологических механизмов регуляции различных функций организма имеет важное значение в понимании хода восстановительных процессов во время и после напряженного мышечного труда.
Раскрывая основные механизмы, обеспечивающие существование целостного организма и его взаимодействие с окружающей средой, физиология позволяет выяснить и исследовать условия и характер изменений деятельности различных органов и систем в процессе онтогенеза человека. Физиология является наукой, осуществляющей системный подход в изучении и анализе многообразных внутри- и межсистемных взаимосвязей сложного человеческого организма и сведение их в конкретные функциональные образования и единую теоретическую картину.
Важно подчеркнуть, что в развитии современных научных физиологических представлений существенная роль принадлежит отечественным исследователям. Знание истории любой науки – необходимая предпосылка для правильного понимания места, роли и значения дисциплины в содержании социально-политического статуса общества, его влияния на эту науку, а также влияние науки и ее представителей на развитие общества. Поэтому рассмотрение исторического пути развития отдельных разделов физиологии, упоминание наиболее ярких ее представителей и анализ естественнонаучной базы, на которой формировались основные понятия и представления этой дисциплины, дают возможность оценить современное состояние предмета и определить его дальнейшие перспективные направления.
Физиологическая наука в России в XVIII–XIX столетиях представлена плеядой блестящих ученых – И. М. Сеченов, Ф. В. Овсянников, А. Я. Данилевский, А. Ф. Самойлов, И. Р. Тарханов, Н. Е. Введенский и др. Но лишь И. М. Сеченову и И. П. Павлову принадлежит заслуга создания новых направлений не только в Российской, но и в мировой физиологии.
Физиологию как самостоятельную дисциплину начали преподавать с 1738 г. в Академическом (позже Санкт-Петербургском) университете. Существенное значение в развитии физиологии принадлежит и основанному в 1755 г. Московскому университету, где в его составе в 1776 г. была открыта кафедра физиологии.
В 1798 г. в Санкт-Петербурге была основана Медико-хирургическая (Военно-медицинская) академия, которая сыграла исключительную роль в развитии физиологии человека. Созданную при ней кафедру физиологии последовательно возглавляли П. А. Загорский, Д. М. Велланский, Н. М. Якубович, И. М. Сеченов, И. Ф. Цион, Ф. В. Овсянников, И. Р. Тарханов, И. П. Павлов, Л. А. Орбели, A.В. Лебединский, М. П. Бресткин и другие выдающиеся представители физиологической науки. За каждым названным именем стоят открытия в физиологии, имеющие мировое значение.
В программу обучения в физкультурных вузах физиология включалась с первых дней их организации. На созданных П. Ф. Лесгафтом в 1896 г. Высших курсах физического образования сразу же был открыт кабинет физиологии, первым руководителем которого являлся академик И. Р. Тарханов. В последующие годы физиологию здесь преподавали Н. П. Кравков, А. А. Вальтер, П. П. Ростовцев, B.Я. Чаговец, А. Г. Гинецинский, А. А. Ухтомский, Л. А. Орбели, И. С. Беритов, А. Н. Крестовников, Г. В. Фольборт и др.
Теоретические предпосылки для возникновения и развития физиологии спорта были созданы фундаментальными работами И. М. Сеченова, И. П. Павлова, Н. Е. Введенского, А. А. Ухтомского, И. С. Бериташвили, К. М. Быкова и других. Однако систематическое изучение физиологических основ физической культуры и спорта началось значительно позже. Особенно большая заслуга в создании этого раздела физиологии принадлежит Л. А. Орбели и его ученику А. Н. Крестовникову, и она неразрывно связана со становлением и развитием Университета физической культуры им. П. Ф. Лесгафта и его кафедры физиологии – первой подобной кафедры среди физкультурных вузов в стране и в мире.
Структура нервного волокна. Проведение нервных импульсов является специализированной функцией нервных волокон, т.е. отростков нервных клеток.
Нервные волокна разделяют намякотные, или миелинизированные, и безмякотные, или немиелинизированные. Мякотные, чувствительные и двигательные волокна входят в состав нервов, снабжающих органы чувств и скелетную мускулатуру; они имеются также в вегетативной нервной системе. Безмякотные волокна у позвоночных животных принадлежат в основном симпатической нервной системе.
Нервы обычно состоят как из мякотных, так и из безмякотных волокон, причем их соотношение в разных нервах различное. Например, во многих кожных нервах преобладают безмякотные нервные волокна. Так, в нервах вегетативной нервной системы, например в блуждающем нерве, количество безмякотных волокон достигает 80—95%. Наоборот, в нервах, иннервирующих скелетные мышцы, имеется лишь относительно небольшое количество безмякотных волокон.
Как показали электронно-микроскопические исследования, мие- линовая оболочка создается в результате того, что миелоцит (шван- новская клетка) многократно обертывает осевой цилиндр (рис. 2.27'), слои ее сливаются, образуя плотный жировой футляр — миелиновую оболочку. Миелиновая оболочка через промежутки равной длины прерывается, оставляя открытыми участки мембраны шириной примерно 1 мкм. Эти участки получили название перехватов Ранвье.

Рис. 2.27. Роль миелоцита (шванновской клетки) в образовании миелиновой оболочки в мякотных нервных волокнах: последовательные стадии спиралеобразного закручивания миелоцита вокруг аксона (I); взаимное расположение миелоцитов и аксонов в безмякотных нервных волокнах (II)
Длина межперехватных участков, покрытых миелиновой оболочкой, примерно пропорциональна диаметру волокна. Так, в нервных волокнах диаметром 10—20 мкм длина промежутка между перехватами составляет 1—2 мм. В наиболее тонких волокнах (диаметром
1—2 мкм) эти участки имеют длину около 0,2 мм. [1]
Безмякотные нервные волокна не имеют миелиновой оболочки, они изолированы друг от друг только шванновскими клетками. В простейшем случае одиночный миелоцит окружает одно безмякот- ное волокно. Часто, однако, в складках миелоцита оказывается несколько тонких безмякотных волокон.
Миелиновая оболочка выполняет двоякую функцию: функцию электрического изолятора и трофическую функцию. Изолирующие свойства миелиновой оболочки связаны с тем, что миелин как вещество липидной природы препятствует прохождению ионов и потому обладает очень высоким сопротивлением. Благодаря существованию миелиновой оболочки возникновение возбуждения в мякот- ных нервных волокнах возможно не на всем протяжении осевого цилиндра, а только в ограниченных участках — перехватах Ранвье. Это имеет важное значение для распространения нервного импульса вдоль волокна.
Трофическая функция миелиновой оболочки, по-видимому, состоит в том, что она принимает участие в процессах регуляции обмена веществ и роста осевого цилиндра.
Прямые электрофизиологические исследования, проведенные Като (1924), а затем Тасаки (1953) на одиночных миелинизированных нервных волокнах лягушки, показали, что потенциалы действия в этих волокнах возникают только в перехватах, а участки между перехватами, покрытые миелином, являются практически невозбудимыми.
В состоянии покоя наружная поверхность возбудимой мембраны всех перехватов Ранвье заряжена положительно. Разности потенциалов между соседними перехватами не существует. В момент возбуждения поверхность мембраны перехвата С становится заряженной электроотрицательно по отношению к поверхности мембраны соседнего перехвата D. Это приводит к возникновению местного (ло 
Рис. 2.28. Сальтаторное распространение возбуждения в мякотном нервном волокне от перехвата к перехвату:
А — немиелинизированное волокно; В — миелинизированное волокно. Стрелками показано направление тока
кального) электрического тока, который идет через окружающую волокно межтканевую жидкость, мембрану и аксоплазму в направлении, показанном на рисунке стрелкой. Выходящий через перехват D ток возбуждает его, вызывая перезарядку мембраны. В перехвате С возбуждение еще продолжается, и он на время становится рефрактерным. Поэтому перехват D способен привести в состояние возбуждения только следующий перехват и т.д.
Синапс, его строение. Виды синапсов. Механизм проведения возбуждения через синапсы. Постсинаптическое и пресинаптическое торможение.
Синапсы - это специализированная структура, которая обеспечивает передачу нервного импульса из нервного волокна на эффекторную клетку - мышечное волокно, нейрон или секреторную клетку.
Синапсы - это места соединения нервного отростка (аксона) одного нейрона с телом или отростком (дендритом, аксоном) другой нервной клетки (прерывистый контакт между нервными клетками).
Все структуры, обеспечивающие передачу сигнала с одной нервной структуры на другую - синапсы.
Значение - передает нервные импульсы с одного нейрона на другой => обеспечивает передачу возбуждения по нервному волокну (распространение сигнала).
Большое количество синапсов обеспечивает большую площадь для передачи информации.
- 1. По локализации: центральные (головной и спинной мозг) и периферические (нервно-мышечный, нейросекреторный синапс вегетативной нервной системы). Центральные синапсы можно в свою очередь разделить на аксо-аксональные, аксо-дендритические (дендритные), аксо-соматические, дендро-дендритические, дендро-соматические и т.п. Согласно Г. Шенсрду, различают реципрокные синапсы, последовательные синапсы и синаптические гломерулы (различным способом соединенные через синапсы клетки).
- 2. По развитию в онтогенезе: стабильные (например, синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе индивидуального развития.
- 3. По конечному эффекту: тормозные и возбуждающие.
- 4. По механизму передачи сигнала: электрические, химические, смешанные.
- 5. Химические синапсы можно классифицировать:
- а) по форме контакта - терминальные (колбообразное соединение) и преходящие (варикозное расширение аксона);
- б) по природе медиатора - холинергические (медиатор - ацетилхолин, АХ), адренергическис (медиатор - норадреналин, НА), дофаминергические (дофамин), ГАМК-ергические (медиатор - гаммааминомасляная кислота), глицинергические, глутаматергические, аспартатсргические, пептидергические (медиатор - пептиды, например, вещество Р), пуринергические (медиатор - АТФ).
Строение синапса:
- 1. Пресинаптическая мембрана - принадлежит нейрону, ОТ которого передается сигнал.
- 2. Синаптическая щель, заполненная жидкостью с высоким содержанием ионов Са.
- 3. Постсинаптическая мембрана - принадлежит клеткам, НА которые передается сигнал.
Между нейронами всегда существует перерыв, заполненный межтканевой жидкостью.
В зависимости от плотности мембран, выделяют:
- - симметричные (с одинаковой плотностью мембран)
- - асимметричные (плотность одной из мембран выше)
Пресинаптическая мембрана покрывает расширение аксона передающего нейрона.
Расширение - синаптическая пуговка/синаптическая бляшка.
На бляшке - синаптические пузырьки (везикуль).
С внутренней стороны пресинаптической мембраны - белковая/гексогональная решетка (необходима для высвобождения медиатора), в которой находится белок - нейрин. Заполнена синаптическими пузырьками, которые содержат медиатор - специальное вещество, участвующее в передаче сигналов.
В состав мембраны пузырьков входит - стенин (белок).
Пузырьки содержат молекулы медиатора (внутри) - вещество, необходимое для передачи сигнала.
Механизм проведения возбуждения через синапсы.
В состоянии покоя в пресинаптической области везикулы постоянно движутся. Они подходят к пресинаптической мембране, выделяя в синаптическую щель медиатор, т. е. в состоянии покоя выделяются несколько квантов медиатора. Он дефилирует через щель, взаимодействует с холинорецепторами. Но т. к. медиатора мало, то потенциал на постсинаптической мембране тоже мал. Это миниатюрные потенциалы с амплитудой в несколько микровольт. Эти потенциалы поддерживают фоновую активность постсинаптической мембраны. Под действием раздражителя возникает потенциал действия нервного волокна. Под действием круговых токов повышается проницаемость мембраны нервного волокна для Са2+, в результате Са2+ поступает внутрь нервного волокна, вызывая упорядоченное движение везикул и, как следствие, увеличение количества медиатора, выделяющегося в синаптическую щель.
На постсинаптической мембране взаимодействие медиатора с рецепторами открывает Nа-каналы, Nа поступает внутрь клетки. Возникает возбуждающий постсинаптический потенциал. Этот потенциал - особая форма возбуждения, которая не распространяется (свойство местного возбуждения). Затем холинэстераза расщепляет медиатор, освобождая рецептор - в результате восстанавливается заряд постсинаптической мембраны.
Постсинаптическое торможение
Торможение - это активный нервный процесс, результатом которого является прекращение или ослабление возбуждения. Торможение вторично относительно процесса возбуждения, так как всегда возникает как следствие возбуждения.
Торможение в ЦНС открыл И.М. Сеченов (1863). В опыте на таламической лягушке он определял латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Было показано, что латентное время рефлекса значительно увеличивается, если на зрительный бугор предварительно положить кристаллик поваренной соли. Открытие И.М. Сеченова послужило толчком для дальнейших исследований торможения в ЦНС. В частности, Ф. Гольц (1870) обнаружил проявления торможения у спинальной лягушки. Он также исследовал латентное время рефлекса. Оказалось, что механическое раздражение кончиков пальцев одной конечности лягушки существенно удлиняет латентный период сгибательного рефлекса другой конечности при погружении ее в раствор кислоты.
Наличие специальных тормозных структур в продолговатом мозге доказал Х. Мегун (1944).
В опытах на кошках при изучении разгибательного рефлекса он установил, что раздражение медиальной части ретикулярной формации продолговатого мозга тормозит рефлекторную активность спинного мозга.
Тонкий анализ тормозных явлений в ЦНС позволил выделить две разновидности торможения:
- 1) постсинаптическое торможение и
- 2) пресинаптическое торможение.
Этот вид торможения открыл Д. Экклс (1952) при регистраци потенциалов мотонейронов спинного мозга у кошки во время сокращения и расслабления мышцы в ходе реализации соответствующих рефлекторных актов. Оказалось, что при рефлекторном расслаблении мышц на мотонейронах регистрируется гиперполяризационный постсинаптический потенциал, уменьшающий возбудимость мотонейрона, угнетающий его способность реагировать на возбуждающие влияния. Поэтому вызванный гиперполяризационный потенциал был назван тормозным постсинаптическим потенциалом (ТПСП).
Пресинаптическое торможение
Пресинаптическое торможение первоначально выявлено в спинном мозге в опыте с регистрацией активности мотонейронов моносинаптической рефлекторной дуги при раздражении антагонистических мышечных нервов.
В очаге деполяризации нарушается процесс распространения возбуждения, следовательно, поступающие импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуде, не обеспечивают выделение медиатора в синаптическую щель в достаточном количестве: нейрон не возбуждается, его функциональное состояние, естественно, остается неизменным.
Разновидности пресинаптического торможения. Обычно выделяют параллельное и латеральное пресинаптическое торможение. Структурная организация этого торможения внешне похожа на такие же варианты постсинаптического торможения. Однако механизм пресинаптического торможения отличается принципиально - возбуждение задерживается не на теле нейрона, а на подступах к нему. В случае постсинаптического торможения заторможен нейрон - снижена возбудимость тела нейрона (обычно вследствие его гиперполяризации).
Локализация в ЦНС пре- и постсинаптического торможения. Оба механизма торможения широко представлены в различных отделах ЦНС: в частности, глицин - медиатор постсинаптического торможения, кроме клеток Реншоу, имеется в ретикулярной формации ствола мозга; ГАМК - в спинном мозге, мозжечке, коре большого мозга. Оба вида торможения могут быть заблокированы: постсинаптическое - стрихнином; пресинаптическое - бику-кулином. Пост- и пресинаптическое торможение блокируется также столбнячным токсином, который нарушает высвобождение тормозных медиаторов.
Потенциалы действия (импульсы возбуждения) обладают способностью распространяться вдоль по нервным и мышечным волокнам. Потенциалы могут быть локальными, они распространяются на небольшие расстояния 1-2 мм с затуханием (декрементом) и импульсными. Импульсные потенциалы распространяются без декремента на значительные расстояния – до нескольких десятков сантиметров. Локальные потенциалы возникают в ответ на действие подпорогового раздражителя, например, на мембране рецепторной клетки. Если локальное возбуждение попадает в участок мембраны, способной генерировать ПД, и амплитуда локального тока достигает критического уровня деполяризации, формируется ПД, который распространяется по всей длине нервного волокна.
Передача информации на большие расстояния в пределах нервной системы осуществляется с помощью нервных импульсов по аксонам нейронов. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциал чувствительных ионных каналов. В зависимости от расположения и концентрации ионных каналов в мембране волокна выделяют два способа проведения нервного импульса.
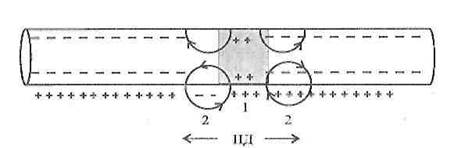
1. Непрерывное проведение нервного импульсаосуществляется в безмиелиновых волокнах, объясняется равномерным распределением потенциал чувствительных ионных каналов, участвующих в генерации ПД.
![]()
Рисунок 3 - Непрерывное распространение ПД в нервном волокне
Возникший ПД обеспечивает открытие потенциал зависимых Na-каналов на соседнем участке мембраны нервного волокна и движение ионов Na + внутрь волокна, что обеспечивает развитие критического уровня деполяризации на соседнем участке нервного волокна и возникновение нового ПД. Непрерывное распространение нервного импульса идет через генерацию новых импульсов по эстафете, когда каждый возникший импульс является раздражителем для соседнего участка нервного волокна и обеспечивает возникновение нового ПД.
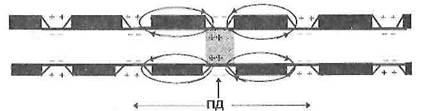
![]()
Рисунок 4 – Сальтаторное распространение ПД в нервных волокнах
2. Сальтаторное проведение нервного импульса (ПД) осуществляется в миелиновых волокнах, так как у них потенциал чувствительные ионные каналы локализованы только в участках мембраны перехватов Ранвье, где их плотность достигает 12 000 на 1 мкм 2 . В области межузловых сегментов, обладающих высокими изолирующими свойствами, потенциал чувствительных каналов нет, вследствие чего мембрана осевого цилиндра там практически невозбудима. Поэтому ПД, возникший в одном перехвате Ранвье распространяется через межузловой сегмент до соседнего перехвата, деполяризует мембрану до критического уровня и вызывает возникновение потенциала действия.
Сальтаторное проведение нервных импульсов является эволюционно более поздним механизмом, возникшим впервые у позвоночных в связи с миелинизацией нервных волокон. Оно имеет два важных преимущества по сравнению с непрерывным механизмом проведения возбуждения:
-более экономично по затрате энергии, так как возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% от площади мембраны волокна, следовательно, требуется меньше энергии для восстановления трансмембранных градиентов ионов натрия и калия;
-возбуждение проводится с большей скоростью (до 120 м/с), чем в безмиелиновых волокнах (0,5-2,0 м/с).
В связи с этими приемуществами миелиновые волокна в нервной системе сформировались там, где необходима наиболее быстрая регуляция функций. Все особенности распространения возбуждения в ЦНС объясняются ее нейронным строением: наличием химических синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей.
Проведение нервных импульсов по волокнам нейронов починяется определенным законам:
Закона 1: нервный импульс распространяется в обе стороны от места раздражения.
Закон 2: проведение нервного импульса по волокну происходит изолировано и не распространяется на параллельные волокна. Объяснение этого закона заключается в том, что аксолемма имеет очень высокое сопротивление и не пропускает петли тока на невозбужденные волокна, расположенные рядом. Изолированное проведение обеспечивает высокую точность регуляторной деятельности ЦНС.
Закон 3: скорость проведения возбуждения по нервному волокну определяется его диаметром. Отсюда следствие: чем толще нервное волокно, тем больше скорость проведения нервного импульса по этому волокну.
Закон 4: нерв сохраняет способность к проведению возбуждения в течение 6-8 часов непрерывного раздражения (закон Н.Е. Введенского, 1883).
Закон 5: действие веществ, блокирующих работу ионных каналов, без нарушения целостности нервного волокна вызывает состояние обратимого парабиоза, (закон парабиоза, Введенский, 1901).
Кроме того, выделяют несколько видов распространения возбуждения:
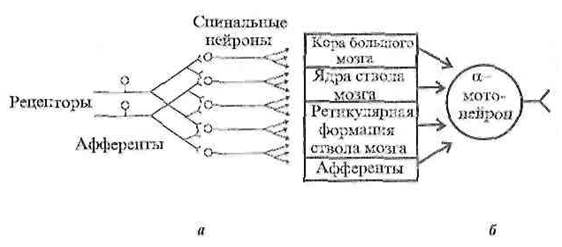
1. Иррадиация (дивергенция) возбуждения в ЦНС, которая объясняется ветвлением аксонов и наличием вставочных нейронов, аксоны которых также ветвятся. Дивергенция расширяет сферу действия каждого нейрона. Один нейрон, посылая импульсы в кору большого мозга, может участвовать в возбуждении до 5000 нейронов.
![]()
Рисунок 4 – Дивергенция афферентных дорсальных корешков на нейроны головного мозга (а – дивергенция, б – конвергенция)
2. Конвергенция возбуждения представляет собой схождение нескольких нервных импульсов, идущих по разным путям к одному и тому же нейрону. Явление конвергенции распространения возбуждения описал Э. Шеррингтон, поэтому явление было названо принцип шеррингтоновской воронки или принцип общего конечного пути. Примером может служить конвергенция возбуждения на спинальном мотонейроне. Мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чиханья и дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг.
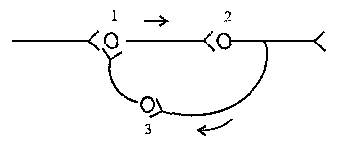
![]()
Рисунок 5 – Циркуляция возбуждения в замкнутых нейронных цепях
3. Циркуляция возбуждения по замкнутым нейронным цепям, возникает в результате замыкания группы нейронов в кольцевую структуру. Циркуляция возбуждения – одна из причин явления последействия. Считают, что циркуляция возбуждения в замкнутых нейронных цепях наиболее вероятный механизм феномена кратковременной памяти. Циркуляция возбуждения возможна в цепи нейронов и в пределах одного нейрона в результате контактов разветвлений его аксона с собственными дендритами и телом.
Литература
1. Смирнов, В.М. Физиология сенсорных систем и высшая нервная деятельность / В.М. Смирнов, С.М. Будылина. - М.: Медицина, 2003. - 304 с.
2. Шульговский, В. В. Основы нейрофизиологии: Учебное пособие для студентов вузов. - М.: Аспект Пресс, 2000. - с. 277.
3. Батуев, А.С. Физиология поведения. Нейрофизиологические закономерности / А.С. Батуев. - Л.: Наука, 1986. - 340 с.
4. Александров, Ю.И. Психофизиология / Ю.И.Александров. - М.: Медицина, 2001. - 230 с.
Данилова, Н.Н. Физиология высшей нервной деятельности / Н. Н. Данилова, А.Л. Крылова. - Ростов н/Д: Феникс, 1999. – 480 с.
[1] ионоселективные каналы и ионные насосы
[2] правило действует и в обратном направлении
[3] 1780 год, профессор анатомии Болонского университета
Читайте также:
