
Передача нервного возбуждения между клетками
Если клеточная мембрана нервного отростка примыкает к мембране другой нервной клетки или к мембране мышечного мышечного волокна то от этого участка через мембрану отростка и мембрану другой клетки будут пассивно распространяться петли тока деполяризуя мембрану соседней клетки. При достижении порогового значения в ней начнётся процесс повышения проводимости к ионам натрия и возникнет потенциал действия. Такое соединение получило название электрического синапса. Для того чтобы передача функционировала более эффективно необходимо увеличить площадь соприкосновения двух мембран, а также плотнее приблизить сами мембраны. Электрическая передача возбуждения имеет определённые преимущества, поскольку ионный ток при такой передаче непосредственно течёт из передающей (пресинаптической) клетки в воспринимающую (постсинаптическую) клетку без каких-либо промежуточных этапов, задержка при проведении возбуждения будет минимальная, и нужно быстро охватить возбуждением нескольких нервных клеток.
В большинстве синапсов используются специальные химические вещества посредники – медиаторы. После возникновения потенциала действия в пресинаптическом окончании мембрана его деполяризована. При этом помимо натриевых и калиевых каналов активируются (открываются) кальциевые каналы и в окончание из внешней среды входят ионы кальция. Повышение внутриклеточной концентрации кальция вызывает выброс медиатора из везикул. Медиатор поступает во внеклеточное пространство и диффундирует к постсинаптической мембране связывается со специальными (рецепторными) участками этой мембраны. В результате происходит активация хемичувствительных ионных каналов и возникает постсинаптический ток, под действием которого развивается потенциал действия мышечной клетки. Так в нервно-мышечном синапсе функционирует ацетилхолин (АцХ) - активируемый канал– интегральный белок мышечной мембраны, состоит из пяти белковых субъединиц, которые образуют трубчатую структуру с общей молекулярной массой около 250000. Рецепторный участок канала состоит из двух белковых субъединиц и располагается на наружной поверхности мембраны. АцХ каналы и связанные с ними АцХ рецепторы сосредоточены в постсинаптической мембране концевой пластинки и плотность их весьма велика – около 10 4 /мкм 2 .
Проведение в химическом синапсе, из-за наличия химического звена, происходит медленнее, чем в электрическом. Однако химическая передача более гибкая, чем электрическая, поскольку с её помощью легко может осуществляться как возбуждающее, так и тормозное действие. Кроме того, при активации постсинаптических каналов химическими агентами может возникать достаточно сильный ток, способный деполяризовать до порогового уровня крупные клетки. В связи с этим при химической передаче тонкие пресинаптические волокна могут возбуждать большие постсинаптические клетки. После окончания действия медиатора он разрушается специальными ферментами. В случае, холинергических синапсов медиатор ацетилхолин гидролизуется холинэстеразой.
Передача возбуждения с нервного волокна осуществляется в синапсе - зоне контакта между нейроном и эффекторными органами (мышца, железа).
В синапсе различают пресинаптическое звено, или пресинапс (концевые утолщения или бляшки аксона), и постсинаптическое звено или постсинапс (участок мембраны дендрита другого нейрона либо мышечной или железистой клетки).
У большинства синапсов синаптическая щель, разделяющая пре- и постсинапс, широкая (10-20 нм) и потому обладает большим сопротивлением, препятствующим непосредственному переходу потенциала действия с пре- на постсинаптическую мембрану. Для этого используется химический посредник - медиатор, вырабатываемый телом нейрона и транспортируемый в окончания аксона, где он накапливается в пузырьках вдоль внутренней поверхности пресинаптической мембраны. Когда под действием нервного импульса наступает ее деполяризация, медиатор выбрасывается в межсинаптическую щель. Обязательным условием для процесса высвобождения медиатора является присутствие ионов Ca 2+ внутри концевого аксонного утолщения.
Далее медиатор связывается с рецепторными белками постсинаптической мембраны, вызывая повышение ее проницаемости для Na + . Поток Na + из синаптической щели через постсинаптическую мембрану ведет к ее деполяризации и генерирует постсинаптический потенциал.
К химическим медиаторам относятся: ацетилхолин, норадреналин, серотонин, аминокислоты - глутаминовая, аспарагиновая, глицин, γ-аминомасляная кислота и другие вещества.
Действие некоторых ядов основано на блокировании синаптической передачи в области нервно-мышечного соединения. Так, действие яда кураре основано на связывании его с рецепторными белками постсинаптической мембраны, что препятствует взаимодействию с ними ацетилхолина. Ботулинический токсин блокирует выход ацетилхолина из пресинапса.
Многие межнейронные синапсы, в отличие от нервно-мышечных, являются тормозными - высвобождающийся медиатор вызывает образование тормозного постсинаптического потенциала, что связано с гиперполяризацией постсинаптической мембраны. Функция тормозных синапсов состоит в подавлении активности нервных клеток.
Синапсы характеризуются рядом функциональных свойств:
- Интервал между поступлением импульса в пресинапс и возникновением постсинаптического потенциала составляет 0,2-0,5 мс (синаптическая задержка).
- Химические синапсы отличаются односторонним проведением: от пре- к постсинапсу. В электрическом синапсе чаще двустороннее проведение.
- Химические синапсы обеспечивают как возбуждение, так*и торможение постсинаптического нейрона, в то время как электрические - только возбуждение.
- Химические синапсы лучше, чем электрические, сохраняют следы предшествующей активности.
- Химические синапсы более чувствительны к изменениям температуры, чем электрические, что имеет существенное значение для нервной системы пойкилотермных (эктотермных) животных.
Количество синапсов на одном нейроне колеблется от нескольких единиц до нескольких тысяч. Это связано с уровнем эволюционного развития животного. Так, 1 г коры головного мозга свинки содержит около 4 · 10 11 синапсов, человека - 1 · 10 18 синапсов.
Взаимосвязи между нервными клетками осуществляются межнейрональ- нымм контактами, или синапсами. Информация в виде серии потенциалов действия поступает от первого (пресинаптического) нейрона на второй (постсинаптический) либо путем формирования локального тока между соседними клетками (электрические синапсы), либо опосредованно химическими веществами — медиаторами, нейротрансмиттерами (химические синапсы), либо с помощью обоих механизмов (смешанные синапсы). Быстрая передача сигналов осуществляется электрическими синапсами, более медленная — химическими.
Типичные синапсы — это образования, сформированные терминалями аксона одного нейрона и дендритами другого (аксодендрипшческие синапсы). Кроме того, существуют аксосомапшческие, аксо-аксональные и дендро-ден- дригпические синапсы. Некоторые ассоциативные нейроны имеют разнообразные синаптические связи. Синапс между аксоном мотонейрона и волокном скелетной мышцы называется двигательной концевой пластинкой или нервно-мышечным соединением.
Электрические синапсы наиболее часто образуются в эмбриональной стадии развития, у взрослого их количество уменьшается. Однако и во взрослом организме значимость электрических синапсов сохраняется для клеток глии и амакринных клеток сетчатки глаза, электрические синапсы можно обнаружить в стволе головного мозга, особенно в нижних оливах, сетчатке глаза, вестибулярных корешках.
Деполяризация пресинаитической мембраны приводит к образованию разности потенциалов с недеполяризованной ностсинаптической мембраной. В результате через каналы, образованные коннексонами, начинается движение положительных ионов но градиенту разности потенциалов в постсинаптическую клетку или движение анионов в обратном направлении. При достижении на постсинаптической мембране суммарной деполяризации пороговой величины возникает потенциал действия. Важно отметить, что в электрическом синапсе ионные токи возникают с минимальной временной задержкой, составляющей К) -0 с, что объясняет высокую синхронизацию ответа даже очень большого числа клеток, соединенных щелевым контактом. Проведение тока через электрический синапс также возможно в обоих направлениях (в противоположность химическому синапсу).
Функциональное состояние электрических синапсов регулируется ионами Са“ + и уровнем мембранного потенциала клеток, что создает условия для влияния на распространение возбуждения вплоть до его прекращения. К особенностям деятельности электрических синапсов следует отнести невозможность прямого переноса возбуждения на отдаленные клетки, так как с возбужденной клеткой непосредственно связаны лишь немногие другие. При этом уровень возбуждения в пресинаитической и ностсинаптической клетках одинаков, затормозить распространение возбуждения невозможно. В связи с этим головной мозг новорожденных и детей раннего возраста, содержащий значительно больше электрических синапсов, чем мозг взрослого, оказывается значительно более возбудимым для электрических процессов: быстро распространяющееся электрическое возбуждение не подвергается тормозной коррекции и почти мгновенно становится генерализованным, что объясняет его особую уязвимость и подверженность развитию пароксизмальной активности.
Химический синапс также передает электрический сигнал от пресинаитической клетки к ностсинаптической, по в нем ионные каналы на постсинаптической мембране открываются или закрываются при помощи химических веществ-переносчиков (медиаторов, нейротрансмиттеров), высвобождающихся из пресинаитической мембраны (рис. 1.6). Изменение возможности

Рис. 1.6. Схема строения и функционирования химического синапса
проведения определенных ионов через постсинаптическую мембрану является основой функционирования химических синапсов. Ионные токи изменяют потенциал ностсинантической мембраны, т.е. вызывают развитие 11 осте инаптического и оте и ц и а л а.
В зависимости от того, проводимость каких ионов изменяется при действии нейротрансмиттера, его эффект может быть тормозящим (гиперполяризация постсинаптической мембраны вследствие дополнительного выходящего тока ионов К + или входящего тока ионов СГ) или возбуждающим (деполяризация ностсинантической мембраны при дополнительном входящем токе ионов Са 2+ или . ! а + ).
Существенным моментом в синаптической передаче является то, что в разных системах межнейрональных связей используются различные медиаторы. В настоящее время известно около 30 химически активных веществ (а цетил холин, дофамин, норадреналин, серотонин, ГАМК и др.), которые играют роль в синаптической передаче импульсов от одной нервной
В синапсе выделяют пресипаптический отросток, содержащий преси- наитические пузырьки (везикулы), и постсинаптическую часть (дендрит, тело клетки, аксон). В иресинаптическом нервном окончании в везикулах аккумулируются нейротрансмиттеры. Синаптические везикулы фиксируются в основном на цитоскелете посредством белков сииапсина, локализованного на цитоплазматической поверхности каждой везикулы, и снектрина, расположенного на волокнах Р-актина цитоскелета. Меньшая часть везикул связана с пресинаптической мембраной посредством белка везикулы сниаптобревииа и белка пресинаптической мембраны синтаксина.
Одна везикула содержит 6000-8000 молекул трансмиттера, что составляет квант трансмиттера, т.е. минимальное количество, освобождающееся в синаптическую щель (рис. 1.7). Когда серия потенциалов действия достигает нервного окончания (пресинаптической мембраны), ионы Са 2+ устремляются внутрь клетки. На связанных с пресинаптической мембраной везикулах ионы Са 2+ связываются с белком везикул синаитотагмином, что вызывает раскрытие мембраны везикул. Параллельно с этим комплекс полипептида синаптофизина сливается с неидентифицированными белками пресинаптической мембраны, что приводит к формированию поры, через которую осуществляется регулируемый экзоцитоз, т.е. секреция нейротрансмиттера в синаптическую щель. Специальные протеины везикул (гаЬЗА) регулируют этот процесс.
Ионы Са 2+ в иресинаптическом окончании активируют Са-кальмоду- лиизависимую протеинкиназу II — фермент, фосфорилирующий синапсин на пресинаптической мембране. Вследствие этого нагруженные трансмиттером везикулы могут освободиться от цитоскелета и переместиться на пре- синаптическую мембрану для осуществления дальнейшего цикла.
Ширина синаптической щели составляет около 20-50 нм. В нее выбрасываются молекулы нейротрансмиттера, локальная концентрация которых сразу после выброса достаточно высока и находится в миллимоляриом диапазоне. Молекулы нейротрансмиттера диффундируют к постсинаптической мембране примерно за 0,1 мс. В постсинаитической мембране встроены протеины, образующие ионные каналы. При попадании молекул нейро- трансмиттера на постсинаптическую мембрану они взаимодействуют с синаптическими рецепторами (белками ионных каналов), изменяя их конформацию и приводя к их открытию. Те рецепторы, ионные каналы которых открываются при непосредственном контакте с лигандом (нейротрансмиттером), называются ионотропными. Те рецепторы, открытие ионных каналов в которых связано с участием других химических процессов, называются м е т а б о т р о п н ы м и. В результате взаимодействия нейротранс-

Рис. 1.7. Основные этапы передачи через химический синапс:
1 — потенциал действия достигает иресинаптического окончания; 2 — деполяризация пресинаитической мембраны приводит к открытию потенциалзависимых Са
+ каналов; 3 — ионы Са 2+ опосредуют слияние везикул с пресинаитической мембраной; 4 — молекулы медиатора высвобождаются в синаптическую щель путем экзо- цитоза; 5 — молекулы медиатора связываются с постсинаитическими рецепторами, активируя ионные каналы; в — происходит изменение проводимости мембраны для ионов и в зависимости от свойств медиатора возникает возбуждающий (деполяризация) или тормозной (гиперполяризация) потенциал иостсинаптической мембраны; 7 — ток ионов распространяется но постсинаптической мембране; 8 — медиа- торные молекулы возвращаются в иресинаптическое окончание путем обратного захвата или .9 — диффундируют во внеклеточную жидкость
миттера с постсинаптическим рецептором возникает возбуждающий постсинаптический потенциал, который пассивно распространяется по всей иостсинаптической мембране. При одновременном возбуждении большого числа синапсов возникает явление суммации ностсинаптического потенциала, что в итоге приводит к генерации потенциала действия, который проводится по аксону постсинаптического нейрона.
Синаптический перенос быстро завершается, и синапс приходит в состояние готовности для нового переноса. Если этот физиологический механизм нарушается, то ответ не возникает и иод влиянием вновь поступающих сигналов возникает блок деполяризации. Механизмом регуляции функционирования синапса является быстрое снижение чувствительности постсинаи- тического рецептора (десенситизация), которая наступает при еще сохраняющихся молекулах нейротрансмиттера. Несмотря на непрерывное связывание нейротрансмиттера с рецептором, конформация каналообразующего белка меняется, ионный канал становится непроницаемым для ионов и синаптический ток прекращается. У многих синапсов десенситизация рецептора может быть длительной (до нескольких минут), пока не произойдут реконфигурация и реактивация канала. Альтернативными физиологическими механизмами прекращения действия трансмиттера, позволяющими избежать длительной десенситизации рецептора, являются быстрое химическое расщепление трансмиттера на неактивные компоненты или его удаление из синаптической щели путем высокоселективного обратного захвата нреси- наптическим окончанием. Характер инактивирующего механизма зависит от типа синапса. Так, ацетилхолин очень быстро гидролизуется ацетилхо- линэстеразой на ацетат и холин. В ЦНС возбуждающие глутаматергические синапсы плотно покрыты отростками аетроцитов, которые активно захватывают нейротрансмиттер из синаптической щели и метаболизируют его.
Синaпс – специализированный контакт между нервными клетками (или нервными и другими возбудимыми клетками), обеспечивающий передачу возбуждения с сохранением его информационной значимости. С помощью синапсов нервные клетки объединяются в нервные сети, которые осуществляют обработку информации. Взаимосвязь между нервной системой и периферическими органами и тканями также осуществляется при помощи синапсов.
По морфологическому принципу синапсы подразделяют на:
нейро-мышечные (аксон нейрона контактирует с мышечной клеткой);
нейро-секреторные (аксон нейрона контактирует с секреторной клеткой);
нейро-нейрональные (аксон нейрона контактирует с другим нейроном):
аксо-соматические (с телом другого нейрона),
аксо-аксональные (с аксоном другого нейрона),
аксо-дендритические (с дендритом другого нейрон).
По способу передачи возбуждения синапсы подразделяют на:
электрические (возбуждение передается при помощи электрического тока);
химические (возбуждение передается при помощи химического вещества):
адренергические (возбуждение передается при помощи норадреналина),
холинергические (возбуждение передается при помощи ацетилхолина),
пептидергические, NO -ергические, пуринергические и т. п.
По физиологическому эффекту синапсы подразделяют на:
возбуждающие (деполяризуют постсинаптическую мембрану и вызывают возбуждение постсинаптической клетки);
тормозные (гиперполяризуют постсинаптическую мембрану и вызывают торможение постсинаптической клетки).
Все синапсы имеют общий план строения (рис. 1).
Конечная часть аксона (синаптическое окончание), подходя к иннервируемой клетке, теряет миелиновую оболочку и образует на конце небольшое утолщение (синаптическую бляшку). Ту часть мембраны аксона, которая контактирует с иннервируемой клеткой, называют пресинаптической мембраной. Синаптическая щель – узкое пространство между пресинаптической мембраной и мембраной иннервируемой клетки, которое является непосредственным продолжением межклеточного пространства. Постсинаптическая мембрана – участок мембраны иннервируемой клетки, контактирующий с пресинаптической мембраной через синаптическую щель.
Рис. 1. Ультраструктура химического и электрического синапса.
Особенности ультраструктуры электрического синапса (см. рис. 1):
узкая (около 5 нм) синаптическая щель;
наличие поперечных канальцев, соединяющих пресинаптическую и постсинаптическую мембрану.
Особенности ультраструктуры химического синапса (см. рис. 1):
широкая (20–50 нм) синаптическая щель;
наличие в синаптической бляшке синаптических пузырьков (везикул), заполненных химическим веществом, при помощи которого передается возбуждение;
в постсинаптической мембране имеются многочисленные хемочувствительные каналы (в возбуждающем синапсе – для Nа+ , в тормозном – для Cl – и К +), но отсутствуют потенциалчувствительные каналы.
Механизм проведения возбуждения аналогичен механизму проведения возбуждения в нервном волокне. Во время развития ПД происходит реверсия заряда пресинаптической мембраны. Электрический ток, возникающий между пресинаптической и постсинаптической мембраной, раздражает постсинаптическую мембрану и вызывает генерацию в ней ПД (рис. 2).
Рис. 2. Передача возбуждения в электрическом синапсе.
Передача возбуждения в химическом синапсе – сложный физиологический процесс, протекающий в несколько этапов. На пресинаптической мембране осуществляется трансформация электрического сигнала в химический, который на постсинаптической мембране снова трансформируется в электрический сигнал.
Медиатором (посредником) называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в химическом синапсе. Некоторые медиаторы (например, ацетилхолин) синтезируются в цитоплазме синаптического окончания, и там же молекулы медиатора депонируются в синаптических пузырьках. Ферменты, необходимые для синтеза медиатора, образуются в теле нейрона и доставляются в синаптическое окончание путем медленного (1–3 мм/сут) аксонного транспорта. Другие медиаторы (пептиды и др.) синтезируются и упаковываются в везикулы в теле нейрона, готовые синаптические пузырьки доставляются в синаптичекую бляшку за счет быстрого (400 мм/сут) аксонного транспорта. Синтез медиатора и образование синаптических пузырьков осуществляется непрерывно.
Секреция медиатора
Содержимое синаптических пузырьков может выбрасываться в синаптическую щель путем экзоцитоза. При опорожнении одного синаптического пузырька в синаптичекую щель выбрасывается порция (квант) медиатора, которая включает около 10000 молекул.
Для активации экзоцитоза необходимы ионы Са++ . В состоянии покоя уровень Са++ в синаптическом окончании низок и выделения медиатора практически не происходит. Приход в синаптическое окончание возбуждения приводит к деполяризации пресинаптической мембраны и открытию потенциалчувствительных Са++ -каналов. Ионы Са++ поступают в цитоплазму синаптического окончания (рис. 3, А,Б) и активируют опорожнение синаптических пузырьков в синаптическую щель (рис. 3, В).
Рис. 3. Передача сигнала в возбуждающем химическом синапсе. А - Д – последовательность процессов при срабатывании химического синапса; Е – деполяризация постсинаптической мембраны (ВПСП).
1 – пресинаптическая мембрана, 2 – синаптическая щель, 3 – постсинаптическая мембрана
Взаимодействие медиатора с рецепторами постсинаптической мембраны
Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны, где связываются с рецепторами хемочувствительных Na+ -каналов (рис. 3, Г). Присоединение медиатора к рецептору приводит к открытию Na+ -каналов, через которые в клетку входят ионы Na+ (рис. 3, Д). В результате входа в клетку положительно заряженных ионов происходит локальная деполяризация постсинаптической мембраны, которую называют возбуждающий постсинаптический потенциал (ВПСП) (рис. 3, Е).
Инактивация медиатора
Ферменты, находящиеся в синаптической щели, разрушают молекулы медиатора. В результате происходит закрытие Na+ -каналов и восстановление МП постсинаптической клетки. Некоторые медиаторы (например, адреналин) не разрушаются ферментами, а удаляются из синаптической щели путем быстрого обратного всасывания (пиноцитоза) в синаптическое окончание.
Генерация ПД
В нейро-мышечном синапсе амплитуда единичного ВПСП достаточно велика. Поэтому для генерации ПД в мышечной клетке достаточно прихода одного нервного импульса. Генерация ПД в мышечной клетке происходит в области, окружающей постсинаптическую мембрану.
В нейро-нейрональном синапсе амплитуда ВПСП значительно меньше и недостаточна для того, чтобы деполяризовать мембрану нейрона до КУД. Поэтому для генерации ПД в нервной клетке требуется возникновение нескольких ВПСП. ВПСП, образовавшиеся в результате срабатывания разных синапсов, электротонически распространяются по мембране клетки, суммируются и генерируют образование ПД в области аксонного холмика. Мембрана нейрона в области аксонного холмика обладает низким электрическим сопротивлением и имеет большое количество потенциалчувствительных Na+ -каналов.
В тормозном химическом синапсе молекулы медиатора, взаимодействуя с рецепторами постсинаптической мембраны, вызывают открытие К+ - и Cl – -хемочувствительных каналов. Вход в клетку Cl– и дополнительная утечка из клетки К+ приводят к гиперполяризации постсинаптической мембраны, которую называют тормозным постсинаптическим потенциалом (ТПСП) . Возникшая гиперполяризация, во-первых, снижает возбудимость клетки. Во-вторых, ТПСП может нейтрализовать возникший в другом месте клетки ВПСП.
Сравнительная характеристика свойств электрических и химических синапсов приведена в табл. 1.
Одностороннее проведение возбуждения в химическом синапсе связано с его функциональной асимметрией: молекулы медиатора выделяются только на пресинаптической мембране, а рецепторы медиатора расположены только на постсинаптической мембране.
Высокая утомляемость химического синапса объясняется истощением запасов медиатора. Утомляемость электрического синапса соответствует утомляемости нервного волокна.
Низкая лабильность химического синапса определяется главным образом периодом рефрактерности хемочувствительных каналов на постсинаптической мембране.
Синаптическая задержка – время от момента возникновения возбуждения в пресинаптической мембране до момента возникновения возбуждения в постсинаптической мембране. Относительно длительное время синаптической задержки в химическом синапсе (0,2–0,7 мс) затрачивается на вход Са++ в синаптическое окончание, экзоцитоз, диффузию медиатора.
Чувствительность синапса к внешним воздействиям определяется характером процессов, протекающих в синапсе при передаче возбуждения. Химические синапсы чувствительны к действию химических веществ, влияющих на синтез и секрецию медиатора, взаимодействие медиатора с рецептором.
Таблица 1.Свойства электрических и химических синапсов

Мы рассмотрели механизмы передачи возбуждения в пределах одной клетки. Что же происходит, когда сигнал доходит до места контакта клетки с другой — нервной, мышечной или иной клеткой организма? Здесь целесообразно сделать небольшое историческое отступление. В начале XX в. крупнейший гистолог С. Рамон-и-Кахаль с помощью светового микроскопа выяснил, что нейроны в нервной ткани окрашиваются как отдельные клетки. Однако многие анатомы продолжали считать, что нервная система представляет собой сеть, в которой цитоплазматическое пространство отростков одной клетки напрямую сообщается с внутриклеточным объемом других клеток. Окончательно этот вопрос был разрешен в 40-х годах с появлением электронного микроскопа, когда были получены четкие данные о наличии мембраны вокруг нервных клеток и их окончаний, а также о существовании особых участков межнейронного взаимодействия, получивших название синапсов.
Прежде чем перейти к описанию структуры и функции синапсов, рассмотрим ситуацию, когда замкнутая клеточная мембрана нервного отростка примыкает к мембране сомы другой нервной клетки или к мембране мышечного волокна. Допустим, в мембране конца нервного отростка (аксона) в результате распространения возбуждения возник потенциал действия. Тогда согласно вышеописанной схеме (см. рис. 2.17) от этого участка через его мембрану и мембрану сомы другой клетки будут пассивно распространяться петли тока (рис. 2.19), деполяризуя мембрану сомы соседней клетки. При
Рис. 2.19. Схема электрического синапса:
стрелкой показано направление распространения потенциала действия в пресинаптическом нервном волокне достижении порогового значения в ней начнется регенеративный процесс повышения проводимости к ионам натрия и возникнет потенциал действия, который может проследовать дальше по отросткам клетки: Такое соединение называется электрический синапс.
Передача через электрический синапс электрического сигнала затруднена несовершенством кабельных свойств клеток. В частности, в том случае, если площадь контакта мембраны нервного волокна и сомы клетки сравнительно небольшая, например контактирует нервный отросток диаметром 5 мкм, общее электрическое сопротивление мембраны этого отростка и участка сомы клетки, к которому он примыкает, составляет миллиарды ом. В то же время сопротивление клеточной мембраны всей сомы клетки будет в несколько десятков раз меньше и сыграет роль короткого замыкания для тока, выходящего из концевой мембраны отростка.
Кроме того, ионный ток будет утекать через пространство между мембранами. Все это, несмотря на высокий фактор надежности — 5. 6 (отношение амплитуды потенциала действия к величине пороговой деполяризации), для потенциала действия чрезвычайно сильно ослабит выходящий из отростка ионный ток и передача возбуждения в этом участке не будет происходить. Для того чтобы передача функционировала более эффективно, необходимо увеличить площадь соприкосновения двух мембран, а также плотнее приблизить сами мембраны. Непосредственная электрическая передача была экспериментально обнаружена сначала на толстых (гигантских) волокнах ракообразных, а затем такие соединения были найдены и у позвоночных животных между нервными клетками в ЦНС, в гладких и сердечных мышцах. Отметим, что электрическая передача возбуждения имеет определенные преимущества. Поскольку ионный ток при такой передаче непосредственно течет из передающей (пресинаптической) клетки в воспринимающую (постсинаптическую) клетку без каких-либо промежуточных этапов, задержка при проведении возбуждения будет минимальная. Таким образом, электрическое проведение более удобно в тех случаях, когда необходимо быстро охватить возбуждением несколько нервных клеток. Вместе с тем в большинстве синапсов нервной и нервно-мышечной систем для передачи сигналов используются специальные химические вещества-посредники — медиаторы. Поскольку следующая часть данной главы посвящена изложению механизмов возбуждения в мышцах, здесь мы остановимся на синаптическом проведении через нервно-мышечный синапс или, как часто называют, через концевую пластинку скелетных мышц позвоночных. О строении и функции синапсов в ЦНС будет подробнее изложено в гл. 12.
Схематически последовательность событий, происходящих при передаче электрического сигнала через химический синапс, можно представить следующим образом (рис. 2.20). После возникно-

Рис. 2.20. Механизм передачи электрического сигнала через химический синапс
вения потенциала действия в пресинаптическом окончании мембрана его деполяризована. При этом помимо натриевых и калиевых каналов активируются (открываются) кальциевые каналы и в окончание из внешней среды входят ионы кальция. Повышение внутриклеточной концентрации кальция вызывает экзоцитоз везикул, наполненных медиатором (выброс из них медиатора). Содержимое везикул поступает во внеклеточное пространство, и часть молекул медиатора, диффундируя к постсинаптической мембране (мембрана мышечной клетки), связывается со специальными (рецепторными) участками этой мембраны. В результате происходит активация ее ионных каналов, связанных с этими молекулами. Движение по каналам соответствующих ионов по их электрохимическим градиентам порождает постсинаптический ток, под действием которого возникает деполяризующий постсинаптический потенциал мышечного волокна. Если этот потенциал превышает пороговый уровень, возникает потенциал действия. Из-за наличия химического звена проведение в химическом синапсе происходит медленнее, чем в электрическом. Однако химическая передача гибче электрической, поскольку с ее помощью легко может осуществляться как возбуждающее, так и тормозное действие. Кроме того, при активации постсинаптических каналов химическими агентами может возникать достаточно сильный ток, способный деполяризовать до порогового уровня крупные клетки. В связи с этим при химической передаче тонкие пресинаптические волокна могут возбуждать большие постсинаптические клетки.
Необходимо отметить, что принятию представленной схемы химической передачи предшествовали в течение шести десятилетий XX в. споры среди ученых о наличии химической передачи и медиаторов в нервно-мышечной системе организма. Первые прямые данные в пользу существования химического медиатора были получены в 20-е годы XX в. О. Леви. Он обнаружил, что при торможении работы сердца лягушки путем раздражения блуждающего нерва из сердца в кровь выделяется вещество, под действием которого частота сокращений сердца другой лягушки также уменьшается. В последующих работах было установлено, что этим веществом — медиатором является ацетилхолин (АцХ). В дальнейшем был обнаружен ряд других медиаторов, накоплено множество данных об их действии и химической природе.
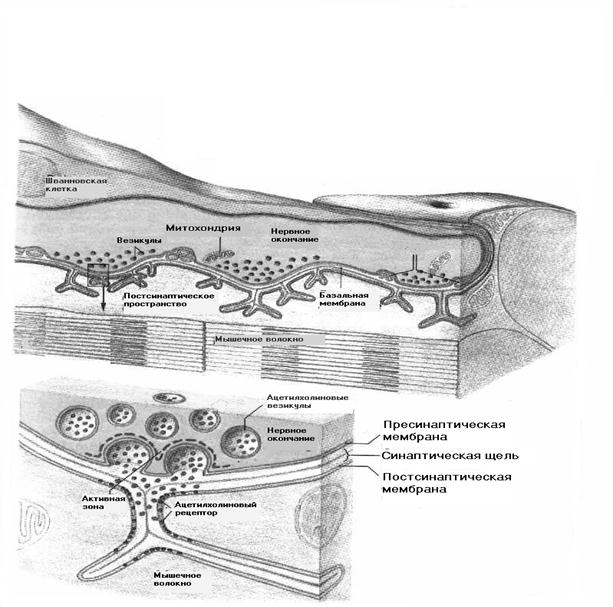
Вернемся вновь к нервно-мышечному синапсу — концевой пластинке. На рисунке 2.21, Л представлено трехмерное изображение концевой пластинки. Она состоит из специализированных участков постсинаптической мембраны, являющейся частью мембраны мышечного волокна, окончания нервного волокна (окончания двигательного нерва — пресинаптической мембраны) и шванновских клеток. От окончания нерва отходят веточки толщиной около 2 мкм, каждая из которых лежит в продольном углублении мембраны мышечного волокна. Мембрана, выстилающая это углубление, с периодичностью 1. 2 мкм образует поперечные субнейрональные складки. В участках нервного окончания, расположенного непосредственно над этими складками, имеются так называемые активные зоны — поперечные участки с несколько утолщенной пресинаптической мембраной, над которыми скапливаются синаптические пузырьки. Пузырьки выделяются из активных зон путем экзоцитоза.
Выделение медиатора из пресинаптического окончания начинается под действием деполяризации этого окончания, вызванной потенциалом действия. Высвободившийся медиатор, в данном случае это АцХ, связывается с рецептором, т. е. со специальной белковой молекулой в постсинаптической мембране концевой пластинки, в результате чего на короткое время открываются каналы, через которые течет ионный ток.
Для выяснения природы ионов, обусловливающих синаптический ток, первоначально изменяли внеклеточные концентрации различных ионов и исследовали влияние этих изменений. Так, было обнаружено, что входящий синаптический ток в концевой пластинке является натриевым ионным током, причем он частично компенсируется выходящим калиевым током. Дальнейшие ис- слёдования показали, что через одни и те же каналы, активируемые в концевой пластинке АцХ, проходят ионы натрия и калия. Значит, эти каналы обладают меньшей избирательностью (селективностью), чем потенциалзависимые натриевые и калиевые каналы при деполяризации мембраны.
Мы уже подробно разбирали механизмы формирования мембранного потенциала в нервных и мышечных клетках и знаем, что

Рис. 2.21. Схема нервно-мышечного синапса (концевой пластинки) и ацетил- хо.1инактивируемого ионного канала:
А — трехмерная реконструкция нервно-мышечного синапса: / — терминальная часть нервного волокна; 2 —синаптические пузырьки; б—синаптические складки; 9— пальцеобразные выросты шванновской клетки; Б— продольный срез участка нервно-мышечного синапса: / — терминаль нервного волокна; 2—синаптические пузырьки; 3— пресинаптичсская мембрана; '/—постсинаптическая мембрана; 5—базальная мембрана; б—синаптическая складка; 7—активная зона; 8, 10 — мышечное волокно; Б а, р, 6, у — белковые субъединицы. На а-белковых субъединицах (одна субъединица не показана) находится участок связывания аиетилхолина (АцХ). Холинрсцегттор взаимодействует с белком цитоскелета (Ц) при изменении мембранной проницаемости для какого-либо иона мембранный потенциал согласно уравнению Голдмана (17) будет смещаться до нового уровня. Если проницаемость для данного иона увеличится, мембранный потенциал сместится ближе к равновесному потенциалу для этого иона. И наоборот, уменьшение проницаемости вызовет смещение в противоположную сторону от равновесного потенциала. В ответ на АцХ каналы концевой пластинки становятся одновременно проницаемы для ионов натрия и калия. При этом возникает смешанный ионный ток, под действием которого потенциал постсинаптической мембраны смещается к новому уровню, так называемому потенциалу реверсии, занимающему промежуточное положение между равновесными потенциалами для этих двух проникающих ионов и равному
10 мВ. В общем случае потенциал реверсии для ионных токов, переносимых двумя ионами, зависит от двух главных факторов: относительной проницаемости активированного канала для проникающих ионов; равновесных потенциалов для этих ионов, зависящих, в свою очередь, от их концентрационных градиентов. Постсинаптические каналы открываются на короткое время (на несколько мс), поскольку АцХ быстро подвергается ферментативному расщеплению ацетилхолинэстеразой (АцХЭ), которая располагается в области субнейрональных складок.
АцХ- активируемый канал. Успехи в совершенствовании методов молекулярной биологии, а также электрофизиологических методов позволили выделить, идентифицировать и изучить АцХ-ак- тивируемый канал — интегральный белок мышечной мембраны. АцХ-активируемый канал состоит из пяти белковых субъединиц, которые образуют трубчатую структуру с общей молекулярной массой около 250000. Канал выступает из мембраны с обеих сторон, причем над наружной ее поверхностью возвышается воронкообразная структура — вход в канал (см. рис. 2.21, В). Рецепторный участок канала состоит из двух белковых субъединиц и располагается на наружной поверхности мембраны. Каждая из белковых субъединиц содержит участок связывания АцХ. Если к этим двум участкам присоединяется АцХ, то канал с высокой вероятностью переходит из закрытого состояния в открытое и через него течет ионный ток.
Читайте также:
