
Порог возбуждения неработающей нервной клетки должен быть
Когда возбуждающий синапс действует на мотонейрон переднего рога, мембрана нейрона становится высокопроницаемой для ионов Na+ в течение 1-2 мсек. За это короткое время достаточное число ионов Na+ быстро диффундируют внутрь постсинаптического мотонейрона, увеличивая его потенциал покоя на несколько милливольт, создавая таким образом ВПСП. Этот потенциал затем медленно убывает на протяжении следующих 15 мсек (это время требуется для того, чтобы избыточные положительные заряды покинули нейрон и восстановился нормальный мембранный потенциал покоя).
Прямо противоположное происходит при развитии ТПСП: тормозной синапс увеличивает проницаемость мембраны к ионам К+ или Сl-, или и к тем и другим, на 1-2 мсек, что ведет к увеличению отрицательного значения потенциала внутри нейрона по сравнению с покоем, т.е. формируется ТПСП. Этот потенциал также исчезает примерно через 15 мсек.
Другие типы медиаторов могут возбуждать или тормозить постсинаптический нейрон на гораздо более длительный период — на сотни миллисекунд или даже несколько секунд, минут или часов. Это особенно справедливо для некоторых пептидных медиаторов.

Возбуждение одной пресинаптической терминали на поверхности нейрона почти никогда не возбуждает его. Это связано с тем, что количество медиатора, выделяемое одной терминалью, обычно достаточно для развития ВПСП, амплитуда которого не более 0,5-1 мВ (вместо 10-20 мВ, что в норме необходимо для достижения порога возбуждения). Однако обычно в одно и то же время стимулируются много пресинаптических терминалей. Даже если эти терминали образуют синапсы в разных частях нейрона, эффекты их влияния могут суммироваться, т.е. складываться друг с другом до тех пор, пока нейрон не возбудится. В основе этого лежат следующие процессы. Как указывалось ранее, изменение потенциала в любой точке внутри сомы вызывает практически такое же изменение потенциала внутри всей сомы.
Это происходит в связи с очень высокой электропроводностью внутри большого тела нейрона. Следовательно, каждый из разряжающихся синхронно возбуждающих синапсов сдвигает общий интрасомальный потенциал на 0,5-1,0 мВ в положительном направлении. Когда ВПСП становится достаточно большим и достигает порога возбуждения, в начальном сегменте аксона спонтанно генерируется потенциал действия. Нижний постсинаптический потенциал на рисунке был вызван стимуляцией 4 синапсов; следующий, более высокий потенциал был вызван стимуляцией 8 синапсов; и наконец, еще более высокий ВПСП был вызван стимуляцией 16 синапсов. В последнем случае был достигнут порог возбуждения, и в аксоне развился потенциал действия.
Эффект суммирования синхронных постсинаптических потенциалов при активации множества терминалей, широко распределенных по поверхности мембраны нейрона, называют пространственной суммацией.
Временная суммация. Каждый раз, когда возбуждается пресинаптическая терминаль, выделяемый ею медиатор открывает мембранные каналы не более чем на 1-2 мсек. Однако изменение постсинаптического потенциала длится до 15 мсек после того, как мембранные каналы в области синапса уже закрылись. Следовательно, повторное открытие тех же каналов может увеличить постсинаптический потенциал до более высокого уровня, и чем больше частота стимуляции, тем выше становится мембранный потенциал. Таким образом, эффекты последовательных разрядов одной пресинаптической терминали при их достаточно высокой частоте могут накладываться один на другой, т.е. суммироваться. Этот тип суммации называют временной суммацией.
Одновременная суммация тормозных и возбуждающих постсинаптических потенциалов. Если ТПСП, увеличивающий отрицательное значение мембранного потенциала, и ВПСП, уменьшающий его отрицательность, развиваются в нейроне одновременно, то эти два эффекта могут полностью или частично нейтрализовать друг друга. Так, если нейрон возбуждается под влиянием ВПСП, тормозной сигнал от другого источника часто может снизить постсинаптический потенциал до уровня ниже порогового, выключая таким образом активность нейрона.
Экспериментальные модели нейронных сетей, строившиеся на основе гипотезы Емельянова-Ярославского, в принципе, обладают свойством активности. Однако предложенная Емельяновым-Ярославским схема работы нейронной сети, так же как и ее модификации, плохо согласуется с принципом устойчивого неравновесия. Возможна и другая схема. Неравновесным состоянием естественно считать состояние повышенной возбудимости. В соответствии с принципом устойчивого неравновесия достигаться максимальное неравновесие (высокая возбудимость) должно не в покое нервной клетки, а наоборот - за счет ее работы, т. е. возбуждений. Порог возбуждения неработающей нервной клетки должен не уменьшаться, а увеличиваться. При каждом срабатывании нейрона его порог возбуждения должен не увеличиваться, а уменьшаться. Оптимальное (неравновесное) состояние нейрона должно соответствовать его повышенной возбудимости, т. е. минимальному значению порога возбуждения.
Рассмотрим на качественном уровне соответствующий вариант активной сети. Экспериментально этот вариант не прорабатывался. Однако достоверность делающихся выводов и предположений во многом основана на опыте экспериментальной работы с рассмотренной модификацией схемы Емельянова-Ярославского, т. е. с моделью трехслойной сети.
В гипотетической модели ( 
-сети), так же как и ранее, предполагается, что нейрон - это пороговый элемент с временной и пространственной суммацией возбуждающих и тормозящих воздействий. Изменяющиеся параметры нейронов - это функциональное состояние, накопленный потенциал возбуждения, порог возбуждения и синаптические проводимости возбуждающих и тормозящих связей. Параметры, отражающие функциональное состояние 
и порог возбуждения 
нейрона, связаны прямой зависимостью 
. (В принципе, о параметре 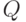
можно было бы не говорить, сводя все к порогу возбудимости, имеющему оптимальное значение и изменяющуюся величину.) Порог возбуждения нейрона увеличивается у несрабатывающего нейрона. При каждом срабатывании порог уменьшается на постоянную величину. Так же изменяется и параметр 
. Возбуждающий потенциал 
образуется алгебраическим суммированием приходящих по связям возбуждающих и тормозящих воздействий. Величины и знаки этих воздействий определяются величинами и знаками синаптических проводимостей связей 
. Проводимости связей изменяются при прохождении по ним сигнала в зависимости от значения параметра
нейрона, на который приходит сигнал. Между одновременно срабатывающими нейронами могут образовываться новые связи. Нейрон срабатывает при 
U" style="display: inline; ">. При срабатывании нейрона возбуждающий потенциал становится равным нулю. При отсутствии воздействий на нейрон накопленный потенциал экспоненциально снижается. Свойства нейронов и связей между ними таковы, что оптимизация в сети возможна лишь при групповом взаимодействии (взаимосодействии) нейронов и их непрерывной или периодической активности.
Начальные качественные предположения об основных общих свойствах нейронных механизмов
-сети состоят в следующем:
- элементом памяти в
![]()
-сети является нейронная структура (нейронный ансамбль) - группа нейронов, объединенных взаимовозбуждающими связями; - эти структуры - ансамбли - формируются в сети при повторяющихся информационных воздействиях, приводящих к одновременному возбуждению группы нейронов;
- на ансамблях нейронов реализуются ассоциации по сходству - весь ансамбль может возбуждаться при приходе внешнего возбуждения только на часть его нейронов;
- между регулярно возбуждающимися последовательно во времени ансамблями устанавливаются возбуждающие связи в прямом направлении времени и тормозящие в обратном. На основе этих связей осуществляется предвидение (предактивация);
- система памяти (модель среды) имеет многоуровневое иерархическое строение, позволяющее формировать обобщения и укрупнения;
- каждый уровень модели среды реализуется в виде трехслойной сети с возвратным торможением;
- свойства нейрона выражаются зависимостями от некоторой переменной величины, соответствующей функциональному состоянию или критерию качества функционирования нейрона и зависящей в каждый момент от интенсивности его срабатываний в течение некоторого предшествовавшего периода (предыстории возбуждений);
- оптимизация по параметру "функциональное состояние" как отдельного нейрона, так и всей сети достигается за счет срабатываний нейронов при их групповом взаимосодействии;
- оптимизация функционального состояния нейронов при приходящих на сеть новых внешних информационных воздействиях достигается за счет структурной перестройки (память);
- при внешних мотивационных воздействиях оптимизация достигается только при снятии этих воздействий в результате целенаправленного поведения;
- решение любых задач в нейронной сети состоит в формировании и гашении очагов возбуждения, мешающих нормальному процессу нормализации общего функционального состояния сети.
Большинство приведенных предположений должны в основном рассматриваться как постулаты, которые определяют свойство активности нейронной сети, направленной на оптимизацию своего функционального состояния. Эти предположения в определенной степени пересекаются с общими исходными свойствами модели Емельянова-Ярославского.
Первично нейроны должны возбуждаться только при внешних активирующих информационных воздействиях. Одновременно возбуждающиеся нейроны образуют ансамбль. Связи между нейронами ансамбля создают синергическое возбуждающее взаимосодействие, обеспечивающее работу (возбуждение) ансамбля и при снятии внешних воздействий. Эта работа должна приводить к достижению оптимальных состояний нейронов ансамбля и их повышенной возбудимости. Заканчиваться работа ансамбля должна при поступлении на него тормозящих воздействий от других работающих ансамблей, а также от элементов, создающих возвратное торможение (схема с возвратным торможением описана выше). Последовательно возбуждающиеся ансамбли должны связываться в цепочки за счет возбуждающих связей в прямом направлении времени и тормозящих в обратном. Это достигается за счет принимаемых в модели свойств связей.
Проводимости связей изменяются при одновременном срабатывании связываемых нейронов. Проводимость возбуждающей связи от нейрона 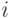
к нейрону 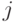
увеличивается, если 
Q_
. Проводимость тормозящей связи от нейрона
к нейрону 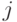
увеличивается, если 
Q_j" style="display: inline; "> и тем сильней, чем больше разность 
. По таким же правилам должны образовываться новые связи между нейронами. В результате будут образовываться ансамбли взаимовозбуждающихся нейронов. Кроме того, будут усиливаться возбуждающие связи, которые идут от ансамбля, начавшего работать раньше, к ансамблю, начавшему работать позже. Также будут усиливаться тормозящие связи, идущие от ансамбля, начавшего работать позже, к ансамблю, начавшему работать раньше.
Если нейроны ансамбля не возбуждаются, то их пороги срабатывания увеличиваются. Одновременно увеличиваются (отклоняются от оптимума) значения параметров
, определяющих функциональные состояния нейронов. Можно сказать, что в
-сети при этом возникает потребность во внешнем информационном запускающем возбуждающем воздействии на ансамбли нейронов. Кроме того, возникает потребность в образовании ансамблей взаимосодействующих нейронов, достигающих оптимальных состояний только в процессе совместной работы и взаимных возбуждений. И наконец, возникает потребность в активации ансамблей от других ансамблей, работа которых при отсутствии внешних информационных воздействий выстраивается в регулярные временные последовательности (предвидение).
Есть еще одно принципиальное отличие от схемы Емельянова-Ярославского. Попытки понять, как устроены и работают нейронные механизмы мозга, должны основываться на том, что мозг развивался в процессе эволюции не для решения своих собственных задач, а для решения задач целого организма. Необходимо ответить на вопрос, как на основе неустойчивости и активности нейронных механизмов решаются не только внутренние, но и внешние функциональные задачи мозга. В первую очередь нужно ответить на вопрос, как осуществляется обучение, то есть построение в мозге активной иерархической модели проблемной среды, и как работает эта активная модель при решении задач восприятия и управления целенаправленным поведением живого организма, а также и при решении абстрактных задач.
Можно представить, что кроме кратковременных информационных возбуждений возможны и длительные возбуждения, мешающие режиму последовательной активации разных нейронных ансамблей, т. е. общей саморегуляции сети. Стабильные фиксированные очаги возбуждения могут соответствовать первичным и вторичным потребностям живого организма. Прекращение возбуждения должно происходить при удовлетворении потребности и снятии соответствующих возбуждающих воздействий. Так связываются поведенческие задачи с задачей оптимизации состояния нейронной сети.
Не менее интересен и вопрос о том, как в мозге возникают, фиксируются и умозрительно решаются задачи абстрактного мышления. В мозге при постановке и решении любых задач, т. е. не только задач поведения, но и умозрительных абстрактных задач, работает принцип доминанты А.А. Ухтомского. При моделировании мышления важно ответить на вопрос, как на нейронном уровне может происходить возникновение, фиксация и снятие доминант или возникновение, поддержание и гашение очагов возбуждения. Возникновение очагов возбуждения, мешающих общей равномерной оптимизации сети, и их гашение соответствует возникновению и решению поведенческих и абстрактных задач мышления, а также поиску информации, необходимой для достройки модели проблемной среды.
В гипотетической полной модели среднее значение параметра
по всей сети должно быть "эмоциональной переменной", то есть должно учитываться в эмоциональном центре при выборе линии поведения наравне с другими эмоциональными переменными, формирующими первичные и вторичные мотивации.
И наконец, важно понять, как в нейронной модели может быть реализован полный отдых в режиме "сна". Этот режим должен приводить к равномерной нормализации функциональных состояний всех нейронов при отсутствии внешних информационных воздействий.
Помимо основных функциональных или рабочих свойств мозг проявляет и специальные свойства и функции, присущие только живому мозгу: это интуиция, озарение, или инсайт, творчество, эмоциональные реакции на музыку, абстрактную живопись (т. е. живопись без смысловой нагрузки), стихотворный ритм, красоту природных явлений. К этому перечню можно добавить еще любопытство, юмор, навязчивую идею, навязчивый мотив. Было бы хорошо, если бы нейронная модель объясняла и такие свойства. Не исключено, что именно активное устойчивое неравновесие нейронов и синергическая активность нейронной сети могут дать ключ к пониманию и этих свойств мозга. Некоторые интерпретации приводятся в дальнейшем изложении. Однако подробная проработка и тем более экспериментальная проверка логики работы активных нейронных сетей, соответствующих принципу устойчивого неравновесия, не проводились.

Перед тем, как говорить о том, каково строение и свойства нейронов, необходимо уточнить, что это такое. Нейрон (рецепторный, эффекторный, вставочный) – функциональная и структурная часть нервной системы, представляющая собой электрически возбудимую клетку. Она отвечает за обработку, хранение, передачу информации химическими и электрическими импульсами.

Весь функционал ЦНС и нервной системы человека зависит от того, насколько хорошо нейроны взаимодействуют друг с другом. Только при совместной работе начинают образовываться сигналы, которые передаются железами, мышцами, клетками организма. Запуск и распространение сигналов происходит посредством ионов, генерирующих электрический заряд, проходимый через нейрон.
Общее число таких клеток в головном мозге человека – около 10 11 , в каждой из которых содержится примерно 10 тыс. синапсов. Если представить, что каждый синапс – это место для хранения информации, то теоретически мозг человека может хранить все данные и знания, которые накоплены человечеством за всю историю его существования.
Физиологические свойства и функции нейронов будут варьироваться в зависимости от того, в какой мозговой структуре они находятся. Объединения нейронов отвечают за регулирование какой-то конкретной функции. Это могут быть самые простые реакции и рефлексы человеческого организма (например, моргание или испуг), а также особо сложный функционал мозговой деятельности.
Особенности строения
Структура включает в себя три основных составляющих:
- Тело. Тело включает в себя нейроплазму, ядро, которое разграничено мембранным веществом. Хромосомы ядра содержат гены, отвечающие за кодировку синтеза белков. Здесь также осуществляется синтез пептидов, которые требуются для обеспечения нормальной работы отростков. Если тело будет повреждено, то в скором времени произойдет и разрушение отростков. При повреждении одного из отростков (при условии сохранения целостности тела) он будет постепенно регенерироваться.
- Дендриты. Образуют дендритное дерево, имеют безграничное число синапсов, сформированных аксонами и дендритами соседних клеток.
- Аксон. Отросток, который, кроме нейронов, не встречается больше ни в одних клетках. Сложно переоценить их значение (например, аксоны ганглиозных клеток ответственны за формирование зрительного нерва).
Классификация нейронов в соответствии с функциональными и морфологическими признаками выглядит следующим образом:
- по числу отростков.
- по типу взаимодействия с другими клетками.
Все нейроны получают грандиозное число электрических импульсов из-за наличия множества синапсов, которые расположены по всей поверхности нейронной структуры. Импульсы также получаются через молекулярные рецепторы ядра. Электрические импульсы передаются разными нейромедиаторами и модуляторами. Поэтому важным функционалом также можно считать способность интеграции полученных сигналов.

Чаще всего сигналы интегрируются и обрабатываются в синапсах, после чего в остальных частях нейронной структуры суммируются постсинаптические потенциалы.
Мозг человека содержит примерно сто миллиардов нейронов. Число будет варьироваться в зависимости от возраста, наличия хронических заболеваний, травм мозговых структур, физической и умственной активности человека.
Развитие и рост нейронов
Современные ученые до сих пор дискутируют на тему деления нервных клеток, т.к. единого мнения по этому вопросу в сфере анатомии на данный момент нет. Многие специалисты в этой области уделяют больше внимания свойствам, а не строению нейронов, что является более важным и актуальным вопросом для современной науки.
Наиболее распространенная версия – развитие нейрона происходит из клетки, деление которой прекращается еще до момента выпуска отростков. Сначала развивается аксон, после чего дендриты.
Зависимо от основного функционала, места расположения и степени активности, нервные клетки развиваются по-разному. Их размеры существенно варьируются в зависимости от места расположения и выполняемых функций.
Основные свойства
Нервные клетки выполняют огромное количество функций. Основные свойства нейрона выглядят следующим образом: возбудимость, проводимость, раздражимость, лабильность, торможение, утомляемость, инертность, регенерация.
Раздражимость считается общей функцией всех нейронов, а также остальных клеток организма. Это их способность давать адекватный ответ на всевозможные раздражения с помощью изменений на биохимическом уровне. Подобные трансформации обычно сопровождаются изменениями ионного равновесия, ослаблением поляризации электрических зарядов в зоне воздействия раздражителя.

Несмотря на то, что раздражимость является общей способностью всех клеток человеческого организма, наиболее выражено она проявляется именно у нейронов, которые связаны с восприятием запаха, вкуса, света и иных подобных раздражителей. Именно процессы раздражимости, протекающие в нервных клетках, запускают другую способность нейронов – возбудимость.
Важнейшее физиологическое свойство нервных клеток, которое заключается в генерировании потенциала действия на раздражитель. Под ним подразумеваются различные изменения, происходящие внутри и снаружи организма человека, которые воспринимаются нервной системой, что и приводит к вызову ответной детекторной реакции. Принято различать два вида раздражителей:
- Физические (получение электрических импульсов, механическое воздействие на разные участки тела, изменение окружающей температуры и температуры тела, световое воздействие, наличие или отсутствие света).
- Химические (изменения на биохимическом уровне, которые считываются нервной системой).
При этом наблюдается разная чувствительность нейронов к раздражителю. Она может быть адекватной и не адекватной. Если в организме человека есть структуры и ткани, которые могут воспринимать конкретного раздражителя, то к нему нервные клетки имеют повышенную чувствительность. Подобные раздражители считаются адекватными (электроимпульсы, медиаторы).

Свойство возбудимости актуально только для нервной и мышечной ткани. Также принято считать, что возбудимостью обладает и ткань желез. Если железа активно работает, то могут отмечаться различные биоэлектрические проявления с ее стороны, потому что она включает в себя клетки разных тканей организма.
Соединительная и эпителиальная ткани не обладают свойством возбудимости. Во время их работы не генерируются потенциалы действия даже в том случае, если происходит непосредственное воздействие раздражителя.
Левое полушарие мозга всегда содержит большее количество нейронов, нежели правое. При этом разница совсем незначительная – от нескольких сотен миллионов до нескольких миллиардов.
Разговаривая о том, каковы свойства нейронов, после возбудимости практически всегда отмечают проводимость. Функция проводника у нервной ткани заключается в особенности проведения возникшего в результате воздействия раздражителя возбуждения. В отличие от возбуждения, функцией проводимости наделены все клетки человеческого тела – это общая способность ткани менять тип своей активной деятельности в условиях воздействия раздражителя.

Повышенная проводимость в нейронных структурах наблюдается при развитии доминантного очага возбуждения. В одном нейроне может происходить конвергенция (объединение сигналов множественных входов, которые исходят от одного источника). Подобное актуально для ретикулярной формации и ряда других систем человеческого организма.
При этом клетки, вне зависимости от структур, в которых они располагаются, могут по-разному реагировать на воздействие раздражителя:
- Изменяется выраженность и выполнение процессов по обмену веществ.
- Изменяется уровень проницаемости мембраны клеток.
- Изменяются биоэлектрические проявления нейронов, двигательная активность ионов.
- Ускоряются процессы развития и деления клеток, повышается выраженность структурных и функциональных реакций.
Выраженность этих изменений также может серьезно варьироваться в зависимости от типа раздражителя, ткани и структуры, в которых находятся нейроны.
Часто можно слышать выражение – нужно предотвращать гибель нервных клеток. Но их гибель запрограммировала природа – за один год человек теряет примерно 1% всех своих нейронов, и никак предупредить подобные процессы нельзя.
Под лабильностью нервных клеток подразумевается скорость течения простейших реакций, которые лежат в основе раздражителя. В обычных условиях, при нормальном развитии всех мозговых структур, у человека отмечается максимально возможная скорость течения. Нейроны, которые различаются электрофизиологическими свойствами и размерами, имеют разные значения лабильности за единицу времени.
В одной нервной клетке лабильность различных структур (аксонной и дендритной частей, тела) будет заметно отличаться. Показатели лабильности нервной клетки определяют с помощью степени ее мембранного потенциала.
Показатели мембранного потенциала должны находиться на определенном уровне, чтобы в нейроне могла получиться наиболее подходящая степень возбудимости и лабильности (зачастую вкупе с ритмической активностью). Только в этом случае нервная клетка сможет в полной мере передать полученную информацию в виде электрических импульсов. Подобные процессы и обуславливают работу нервной системы в целом, а также гарантируют нормальное протекание и формирование всех необходимых реакций.
В спинном мозге предельный уровень ритмической активности нервных клеток может достигать значения в 100 импульсов в секунду, что соответствует наиболее оптимальным значениям мембранного потенциала. В обычных условиях данные значения редко превышают уровень в 40-70 импульсов в секунду.

Существенное превышение показателей наблюдается при характерных выраженных реакциях, поступающих со стороны главных отделов ЦНС, мозговых структуры, коры. Частота разрядов при определенных условиях может достигать значений в 250-300 импульсов в секунду, но подобные процессы развиваются крайне редко. Также они являются кратковременными – их быстро сменяют замедленные ритмы активности.
Наиболее высокие показатели частоты разрядов обычно наблюдаются в нервных клетках спинного мозга. В возникающих в результате выраженного воздействия раздражителя очагах начальных реакций частота разрядов может составлять 700-1000 импульсов в секунду. Протекание подобных процессов в нейронных структурах является необходимостью, чтобы клетки спинного мозга могли резко и быстро воздействовать на мотонейроны. Спустя небольшой промежуток времени частота разрядов существенно снижается.
Нейроны существенно различаются по размеру (в зависимости от места расположения и других факторов). Размеры могут варьироваться от 5 до 100 мкм.
С точки зрения физиологии человека торможение, как ни странно, является одним из наиболее активных процессов, протекающих в нейронных структурах. Особенности строения и свойств нейронов подразумевают, что торможение вызывается возбуждением. Процессы торможения проявляются в снижении активности или предупреждении вторичной волны возбуждения.
Способность нервных клеток к торможению совместно с функцией возбуждения позволяет обеспечить нормальную работу отдельных органов, систем, тканей организма, а также всего человеческого тела в целом. Одна из наиболее важных характеристик процессов торможения в нейронах – обеспечение защитной (охранной) функции, что актуально для клеток, располагающихся в коре головного мозга. За счет процессов торможения также обеспечивается защита ЦНС от чрезмерного перевозбуждения. Если они нарушены, у человека проявляются негативные психоэмоциональные черты и отклонения.

Важной функцией торможения также является прямое взаимодействие с возбуждением, что позволяет анализировать и синтезировать в центральной нервной системе полученные электрические импульсы. Это помогает правильно согласовывать деятельность и функции всех систем, тканей и органов человеческого тела, а также адекватно контактировать с окружающей средой. Данную функцию также принято называть координационной.
Несмотря на то, что нейроны имеют удивительно малые размеры, современные технологии позволяют ученым провести измерение активности каждого найденного нейрона. Подобные процедуры зачастую проводятся для диагностики различных заболеваний (например, эпилепсии).
К общим признакам всех нейронов относится их способность к физиологической и репаративной регенерации. В нервных клетках она подразумевает протекание следующих процессов:
- Частичное увеличение количества хромосом в ядре.
- Восстановление синапсов (если они были повреждены).
- Развитие и возвращение в обычное состояние отростков (при их повреждении).
- Обновление метаболических и химических компонентов нервных клеток в процессе протекания внутриклеточного обмена веществ.
Если нервная ткань будет повреждена, то в зоне поражения сразу начнет развиваться нейроглия. Это невыраженная дифференцированная ткань, которая делится митозом.
В случае получения повреждений, которые нарушили целостность нервных волокон, происходит распадение периферических частей на отдельные части миелиновых оболочек и осевых цилиндров. Если отсутствуют воспалительные процессы, рубцы соединительной ткани, то есть высокая вероятность восстановления иннервации нервных тканей. Отростки нейронов регенерируются довольно быстро – 2-3 мм за 24 часа.
Вопреки распространенному мнению, нейроны вполне могут восстанавливаться – их генерирование происходит сразу в трех частях организма человека.
Читайте также:
