
Симпатическая нервная система это адренорецепторы
а) Ганглионарная передача нервных импульсов. Преганглионарные нейроны симпатической и парасимпатической систем — холинергические: при образовании аксодендритических синапсов с ганглионарными клетками из этих нейронов высвобождается ацетилхолин (АХ). Рецепторы на поверхности ганглионарных клеток называют никотиновыми, так как их возбуждение может происходить при местном действии никотина.
б) Передача нервных импульсов в нейроэффекторном синапсе. Постганглионарные нервные волокна симпатической и парасимпатической систем образуют нейроэффекторные соединения (синапсы)с эффекторными тканями (тканями-мишенями). За высвобождение нейромедиаторов отвечают многочисленные пресинаптические утолщения, расположенные по ходу нервных волокон.
Главный нейромедиатор в симпатических нейроэффекторных соединениях — норадреналин (норэпинефрин), который высвобождается из гранулярных везикул. Постганглионарные симпатические волокна — преимущественно адренергические; исключение составляют холинергические волокна, отвечающие за симпатическую иннервацию расположенных по всему телу эккриновых потовых желез. Главный нейромедиатор в парасимпатических нейроэффекторных соединениях — ацетилхолин (АХ). Постганглионарные парасимпатические волокна преимущественно холинергические.
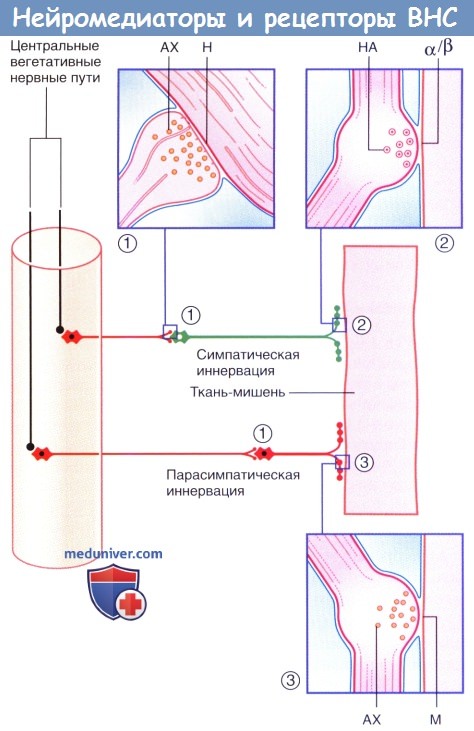
Нейромедиаторы и рецепторы вегетативной нервной системы.
(1) Аксодендритические синапсы с никотиновыми рецепторами.
(2) Нейроэффекторные синапсы с адренергическими рецепторами.
(3) Нейроэффекторные синапсы с мускариновыми рецепторами. Ганглионарные нейроны и постганглионарные волокна выделены красным цветом.
АХ — ацетилхолин; М — мускариновые рецепторы; Н — никотиновые рецепторы; НА — норадреналин.
в) Рецепторы в нейрозффекторном синапсе. Физиологические эффекты вегетативной нервной системы (ВНС) зависят от вида рецепторов на постсинаптической мембране (плазматической мембране эффекторных клеток). На высвобождение нейромедиаторов также влияют рецепторы пресинаптической мембраны (аксолеммы).
- Синаптические рецепторы симпатической системы (адренорецепторы). Для норадреналина существуют два вида а-адренорецепторов и два вида β-адренорецепторов.
1. Постсинаптические α1-адренорецеторы, активация которых вызывает сокращение гладких мышц мелких периферических артерий и крупных артериол, дилататора зрачка, а также мышц семявыносящего протока, сфинктеров ЖКТ и шейки мочевого пузыря.
2. Пресинаптические α2-адренорецеторы локализуются как на симпатических, так и на парасимпатических нервных окончаниях и ингибируют в них высвобождение нейромедиаторов. Пресинаптические α2-адренорецеторы симпатической системы называют ауторецепторами.
3. Постсинаптические адренорецепторы, возбуждение которых приводит к увеличению пейсмекерной активности клеток сердца, а также к повышению силы сердечных (желудочковых) сокращений. При резком падении артериального давления за счет активации симпатических β1-адренорецепторов юкстагломерулярных клеток почек происходит секреция ренина, который, в свою очередь, вызывает секрецию мощного вазоконстриктора ангиотензина II.
4. β2-Адренорецепторы, которые реагируют как на норадреналин, так и на адреналин (эпинефрин).
Активация постсинаптических β2-адренорецепторов вызывает расслабление гладких мышц, наиболее ярко выраженное в трахеобронхиальном дереве и мышцах глаза, участвующих в аккомодации. Часть постсинаптических β2-адренорецепторов располагается на поверхности гепатоцитов. При возрастании энергетических затрат организма их активация вызывает распад гликогена, необходимый для поддержания уровня глюкозы в крови.
Пресинаптические β2-адренорецепторы адренергических нервных окончаний отвечают за высвобождение норадреналина.
В симпатических нервных окончаниях большая часть высвобождаемого норадреналина подвергается обратному захвату моноаминным ферментным насосом. После обратного захвата часть норадреналина разрушается митохондриальным ферментом — моноаминоксидазой (МАО). Влияние лекарственных средств на симпатическую систему подробно рассмотрено в блоке клинической информации ниже.
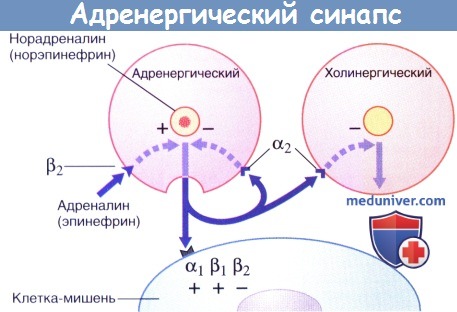
Передача возбуждения в адренергическом нейроэффекторном синапсе.
Секреция норадреналина стимулируется адреналином и ингибируется путем активации пресинаптических α2-рецепторов (данные рецепторы активируются и на пресинаптической мембране близлежащих парасимпатических синапсов, где также ингибируют секрецию).
- Синаптические рецепторы парасимпатической системы. Парасимпатические синаптические рецепторы называют мускариновыми, так как их активацию, помимо АХ, может вызывать другое химическое вещество — мускарин. Стимуляция парасимпатической системы приводит к следующим М-холинергическим эффектам.
• Замедление (в ответ на стимуляцию блуждающего нерва) частоты сердечных сокращений, а также уменьшение силы желудочковых сокращений.
• Сокращение гладких мышц, которое обусловливает такие процессы, как перистальтика кишечника, опустошение мочевого пузыря, а также рефлекс аккомодации при взгляде на близкое расстояние.
• Секреция желез.
Помимо представленных выше эффектов, пресинаптические мускариновые рецепторы симпатических нервных окончаний также отвечают за ингибирование высвобождения норадреналина.
Влияние лекарственных средств на парасимпатическую систему подробно рассмотрено в Блоке клинической информации. Лекарственные средства, обладающие мускариноподобным действием, называют холинергическими (холиномиметическими). Лекарственные средства, блокирующие доступ АХ к постсинаптической мембране, называют антихолинергическими (холинолитическими).
Главный фактор, который необходимо учитывать при назначении препаратов, активирующих или подавляющих деятельность симпатической и парасимпатической систем,— наличие рецепторов к этим лекарственным веществам α-, β- и мускариновых) в ЦНС. Так, в частности, применение данных лекарственных средств в психиатрии обусловлено их действием на центральные, а не на периферические рецепторы.
Расположение парасимпатических интрамуральных ганглиев в сердце. 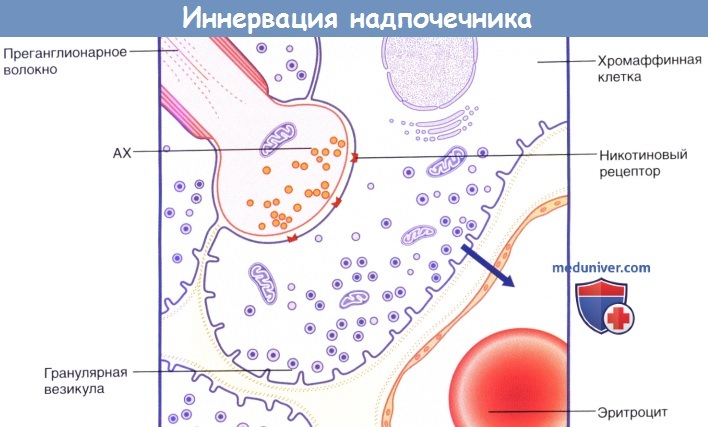
Синаптическое соединение преганглионарного волокна грудного внутренностного нерва и хромаффинной клетки мозгового вещества надпочечников.
Активация никотиновых рецепторов ацетилхолином (АХ).
8% клеток содержат крупные гранулярные везикулы (представлены на рисунке), высвобождающие адреналин; его секрекция в капиллярное русло обозначена стрелкой.
20% клеток содержат маленькие гранулярные везикулы, высвобождающие норадреналин. 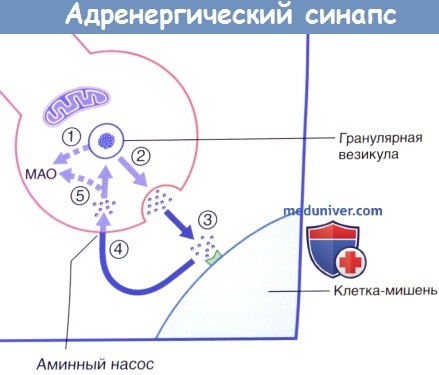
Высвобождение и обратный захват нейромедиатора в адренергическом нервном окончании.
МАО — моноаминоксидаза. 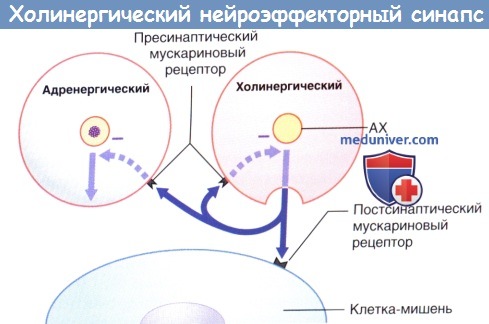
Передача возбуждения в холинергическом нейроэффекторном синапсе.
Избыточная секреция ацетилхолина (АХ) ингибируется путем активации пресинаптических мускариновых рецепторов (данные рецепторы активируются и на пресинаптической мембране близлежащих симпатических синапсов, где также ингибируют секрецию).
г) Другие типы нейронов. И в симпатическом, и в парасимпатическом отделах ВНС часть нейронов представлена неадренергическими, нехолинергическими нейронами (NANC-нейронами). Находящиеся в симпатических ганглиях небольшие вставочные нейроны отвечают за высвобождение дофамина — предшественника норадреналина. Часть дофамина выделяется в капиллярное русло, другая часть связывается с дофаминовыми рецепторами на основных (адренергических) нейронах, оказывая на них слабое тормозное действие.
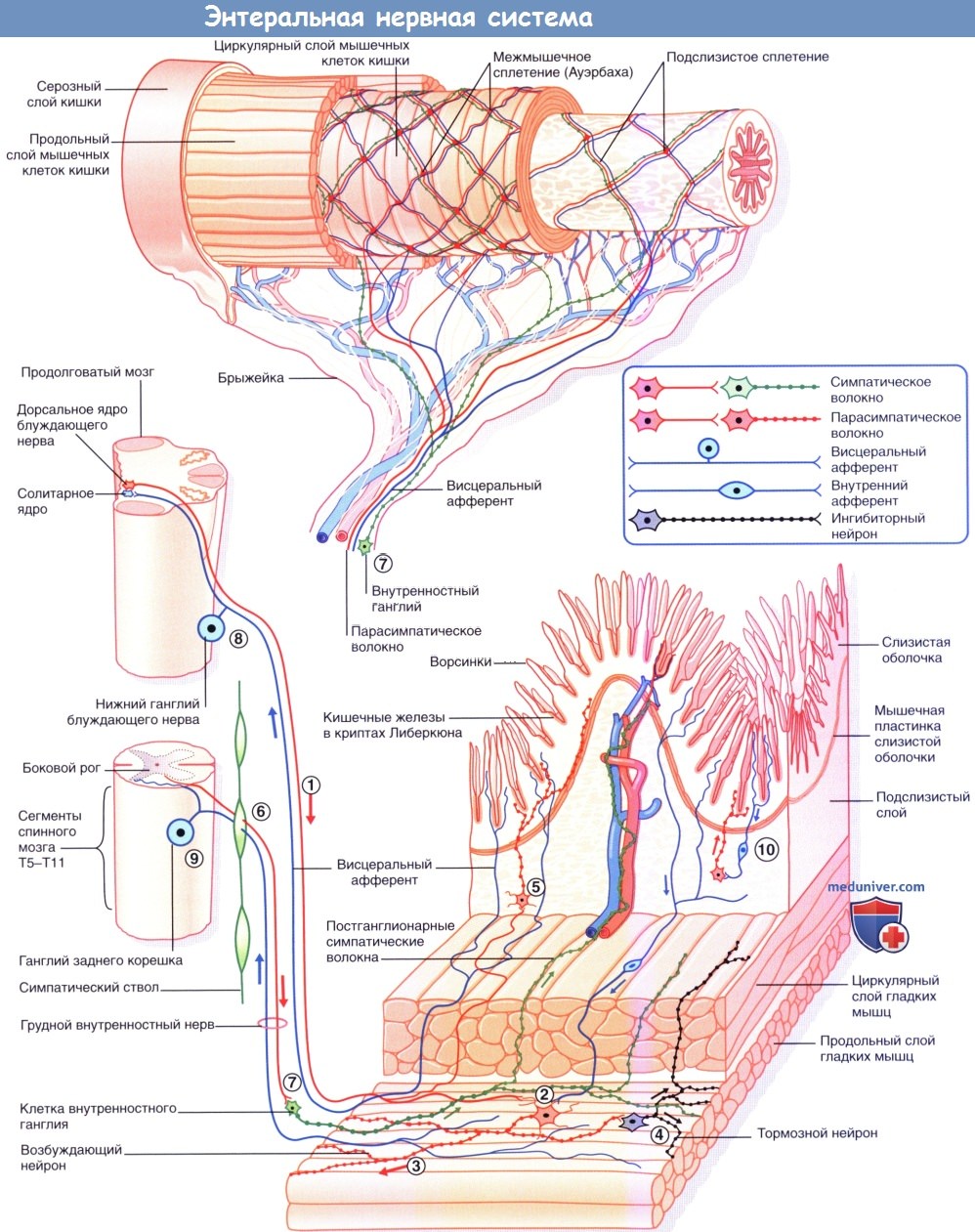
Самое большое количество NANC-нейронов находится в области ганглионарных клеток стенки ЖКТ и в тазовых ганглиях. Эти нейроны содержат более 50 разнообразных белковых субстанций, находящихся в несвязанном состоянии или в различных комбинациях. Функция большей части этих веществ — регуляторная: действуя на пресинаптическую или постсинаптическую мембрану, они влияют на продолжительность работы основных нейромедиаторов. Другие белковые субстанции (вещества, выделяющиеся совместно с ацетилхолином (АХ)) представляют собой комедиаторы (сопутствующие медиаторы).
Редактор: Искандер Милевски. Дата публикации: 14.11.2018
Симпатическая часть вегетативной нервной системы состоит из центрального и периферического отделов. Центральный отдел образуют клетки боковых рогов спинного мозга на уровне всех его грудных и верхних трех поясничных сегментов. Периферический отдел представлен нервными волокнами и симпатическими нервными узлами (ганглиями). Нервные узлы подразделяются на две группы:
околопозвоночные (паравертебральные), расположенные двумя цепочками по бокам от позвоночника и образующие правый и левый симпатические стволы, и предпозвоночные (превертебральные) — узлы периферических нервных сплетений, лежащие в грудной и брюшной полостях
Основным медиатором симпатических синапсов является норадреналин и такие синапсы называютадренергическими.Рецепторы, связывающие адренергический медиатор получили названиеадренорецепторов.Различают два типа адренорецепторов — альфаи бета,каждый из которых делят на два подтипа — 1 и 2. Небольшая часть симпатических синапсов использует медиатор ацетилхолин и такие синапсы называют холинергическими,а рецепторы — холинорецепторами.Холинергические синапсы симпатической нервной системы обнаружены в потовых железах. В адренергических синапсах кроме норадреналина в существенно меньших количествах содержатся адреналин и дофамин, также относящиеся к катехоламинам, поэтому медиаторное вещество в виде смеси трех соединений раньше называли симпатином.
Норадреналин (НА), из группы катехоламинов, производное аминокислоты тирозина Постганглионарные окончаниясимпатическойвегетативной нервной системы. α1- , α2-, β1-, β2-адренорецепторы (все метаботропного типа) Возбуждение α1- и β1- рецепторов оказывает возбуждающее действие, а α2- и β2-рецепторв - тормозящее.
Отсюда - набор симпатических реакций:
1) усиление сокращений сердца,
2) сужение сосудов мышц,
3) расширение бронхов,
4) ослабленние перистальтики кишечника,
5) расширение зрачков.
Обычно в одном органе содержится несколько типов адренорецепторов. При этом α-рецепторы преобладают в сосудах кожи и сфинктерах ЖКТ, β1-рецепторы - в сердце, β2-рецепторы - в бронхах, кишечнике, матке, сосудах скелетных мышц.
6. Характеристика парасимпатического отдела вегетативной нервной системы его медиаторы
Центрами парасимпатического отдела автономной нервной системы являются ядра, находящиеся в среднем мозге (IIIпара черепно-мозговых нервов), продолговатом мозге (VII, IXи Х пары черепно-мозговых нервов) и крестцовом отделе спинного мозга (ядра тазовых внутренних нервов). От среднего мозга отходят Преганглионарные волокна парасимпатических нервов, которые входят в состав глазодвигательного нерва. Из продолговатого мозга выходят Преганглионарные волокна, идущие в составе лицевого, языкоглоточного и блуждающих нервов. От крестцового отдела спинного мозга отходят Преганглионарные парасимпатические волокна, которые входят в состав тазового нерва.
При возбуждении парасимпатических нервов тормозится работа сердца отрицательные хроно-, ино-, дромо- и батмотропное действия),повышается тонус гладкой мускулатуры бронхов, в результате чего уменьшается их просвет, сужается зрачок, стимулируются процессы пищеварения (моторика и секреция), обеспечивая тем самым восстановление уровня питательных веществ в организме, происходит опорожнение желчного пузыря, мочевого пузыря, прямой кишки. Действие парасимпатической нервной системы направлено на восстановление и поддержание постоянства состава внутренней среды организма, нарушенного в результате возбуждения симпатической нервной системы. Парасимпатическая нервная система выполняет в организметрофотропную функцию.
К этому отделу относятся интрамуральные системы всех полых висцеральных органов, обладающих собственной автоматической двигательной активностью: сердце, бронхи, мочевой пузырь, пищеварительный тракт, матка, желчный пузырь и желчные пути.
Внутриорганный отдел имеет все звенья рефлекторной дуги: афферентный, вставочный и эфферентный нейроны, которые полностью находятся в органе и нервных сплетениях внутренних органов (например, ауэрбаховском и мейснеровском). Преганглионарные волокна выделяютацетилхолининорадреналин,постганглионарные —АТФиаденозин, ацетилхолин, норадреналин, серотонип, дофамин, адреналин, гистамини т.д. Главная роль принадлежит АТФ и аденозину.
Ацетилхолин (АХ), низкомолекулярное вещество с зарядом +1
Дата добавления: 2019-11-25 ; просмотров: 465 ;
Автономная (вегетативная, висцеральная) нервная система, как уже указывалось осуществляет регуляцию функций без участия или с минимальным участием ЦНС т.е. бессознательно. Дуга вегетативного рефлекса в своей афферентной (сенсорной) части существенно не отличается от дуги соматического рефлекса, более того данное сенсорное волокно может вызывать как соматический, так и вегетативный рефлексы. Но в отличие от моносинаптических рефлексов растяжения даже простейшая дуга спинальных вегетативных рефлексов содержат вставочные интернейроны. Дуги этих рефлексов различаются также локализацией эффекторных нейронов. Эффекторные нейроны вегетативной системы, обычно называемые постганглионарными нейронами, расположены полностью за пределами ЦНС, тогда как соматические мотонейроны лежат в сером веществе спинного мозга. Промежуточные нейроны (интернейроны) дуги вегетативного рефлекса называют преганглионарными нейронами. Вегетативную нервную систему подразделяют на симпатический (грудопоясничный)и парасимпатический (черепнокресцовый) отделы.
Преганглионарные нейроны в симпатическом отделевегетативной нервной системы располагаются в грудном и поясничном отделах спинного мозга и образуют н-холинэргические синапсы (синапсы у которых на постсинаптической мембране имеются н-холинорецепторы) с постганглионарными нейронами в двух цепочках вегетативных ганглиев, расположенных по обе стороны позвоночника от основания черепа до крестца, а также в автономных ганглиях, расположенных на значительном удалении от спинного мозга и вдали от иннервируемых органов. К числу наиболее крупных автономных (превертебральных) ганглиев относятся чревный и краниальный, которые вместе с отходящими нервами образуют самое крупное из автономных нервных сплетений - солнечное сплетение. Цепочки вегетативных ганглиев называются соответственно правым и левым симпатическими стволами. От них и автономных ганглиев идут к иннервируемым органам довольно длинные постганглионарные аксоны, группирующиеся в отдельные нервы, каждый из которых имеет свое название.
Преганглионарные нейроны парасимпатического отдела находятся в черепном и крестцовом отделах ЦНС. Их аксоны образуют н-холинэргические синапсы с постганглионарными нейронами, расположенными не в отдельных ганглиях, а в самих висцеральных органах или около них. Поэтому в парасимпатическом отделе отдельные нервы (наиболее известный блуждающий нерв) образованы длинными преганглионарными аксонами.
Внутренние органы - сердце, пищеварительный тракт, кровеносные сосуды, железы и т.д. получают двойную иннервацию - от симпатического и парасимпатического отделов вегетативной нервной системы, которые оказывают обычно противоположное действие на функции этих органов (висцеральные функции). Так например, деятельность сердца угнетается под влиянием блуждающего нерва (парасимпатическая иннервация) и усиливается в результате импульсации, приходящей по симпатическим нервам. На пищеварительный тракт (моторику кишечника и секрецию пищеварительных соков) парасимпатическая и симпатическая системы оказывают взаимно обратный эффект.
Симпатическая нервная система имеет тенденцию возбуждаться как единое целое, вызывая генерализованные реакции по всему телу. Она активируется в условиях испуга, нападения или бегства, а также в процессе интенсивной физической нагрузки. Симптомы ее хорошо известны: расширенные зрачки, сухость во рту, колотящееся сердце, потение, сильные переживания. Парасимпатическая система, наоборот, отвечает за восстановление ресурсов, уменьшает частоту сердцебиений стимулирует деятельность пищеварительной системы, В общем, парасимпатическая система является более избирательной в своей деятельности. Есть существенная выгода в том, что зрачок может сокращаться при ярком освещении, а хрусталик глаза — аккомодироваться для рассматривания соседствующих объектов избирательно, без сопутствующих и весьма несвоевременных сокращений мочевого пузыря либо еще менее уместных парасимпатических эффектов (в половой сфере).
Различия в воздействии симпатической и парасимпатической систем на висцеральные функции обусловлено тем, что их нервные окончания выделяют разные медиаторы. Постганглионарные волокна парасимпатической системы выделяют ацетилхолин, который воздействует, главным образом, на мускариновые рецепторы. Эти рецепторы опосредуют тормозные эффекты в одних тканях и возбудительные в других. Постганглионарные волокна симпатической системыв качестве медиатора выделяют катехоламины (норадреналин) и образуют контакты с норадренэргическими рецепторами двух типов: альфа и бета-рецепторами. Альфа-рецепторы избирательно блокируются феноксибензамином, тогда как бета-рецепторы блокируются пропранололом. Активация альфа-рецепторов ведет к повышению внутриклеточного уровня Са 2+ , а бета-рецепторы регулируют активность связанного с мембраной фермента аденилатциклазы, ответственного за образование циклического аденозинмонофосфата (цАМФ). Следует иметь в виду, что постганглионарные участки выделения медиаторов в тканях внутренних органов и постсинаптические рецепторные зоны не образуют четко ограниченных областей подобно тому как это имеет место в таких классических нейроэффекторных синапсах, как например нервномышечные синапсы.
В настоящее время установлено, что альфа-адренорецепторы делятся на несколько подтипов, в частности α1-рецепторы и α2-рецепторы. Описанные выше свойства характерны для α1-рецепторов на постсинаптической мембране, в то время как α2-рецепторы чаще локализованы на пресинаптической мембране и называются ауторецепторами. Норадреналин не только активирует рецепторы на постсинаптических клетках-мишенях, но также действует на сами пресинаптические норадренергические нервные окончания через α2-адренорецепторы. Активация ауторецепторов ослабляет выделение медиатора, снижая вероятность открывания кальциевых каналов посредством G-белков. Активный βγ-комплекс G-белков связывается с кальциевыми каналами, что приводит к уменьшению входа кальция и освобождения медиатора. Такой тормозный эффект является быстрым и локальным механизмом для регуляции выброса норадреналина в симпатической системе. Однако, α2-рецепторы обнаружены не только в вегетативной нервной системе, но и головном мозге, где их специфическим агонистом является клофелин. Это гипотензивный препарат, обладает седативным и снотворным эффектами. Действие его может быть значительно потенцировано (усилено) снотворными, психотропными средствами и алкогольными напитками.
В ВНС выделяют симпатический, парасимпатический и метасимпатический отделы. Симпатический и парасимпатический отделы имеют центральное представительство и периферическое звено. В полых внутренних органах, обладающих собственной ритмической моторной активностью (сердце, желудок, кишечник, мочевой пузырь и др.), расположен третий (внутриорганный) метасимпатический отдел.
Симпатический отдел. Симпатические центры образованы преганглионарными нейронами боковых столбов спинного мозга на протяжении шейных, грудных и поясничных сегментов С8—L3 (спиноцилиарный центр — С8—Т2, слюнной центр — Т2—Т4, сердечный центр — Т,—Т5, сосуды и потовые железы — С8—L3 и др.). Они активируются афферентными стимулами, поступающими в спинной мозг, а также влияниями вышележащих структур головного мозга (особенно нейронов ретикулярной формации). Областью иннервации симпатических центров являются практически все органы и ткани (рис. 2.2.4).
Преганглионарные волокна, выходящие из симпатических центров, образуют миелиновые аксоны преганглионарных нейронов (тип В, скорость проведения возбуждения около 10 м/с). Потенциалы действия в этих волокнах имеют длительную следовую деполяризацию. Это свидетельствует о повышенной возбудимости нейрона после спайка потенциала действия (ПД). Преганглионарные волокна поступают в нервные узлы (ганглии).
Ганглии симпатической нервной системы расположены около позвоночника (паравертебральные ганглии) и впереди позвоночника (превертебральные ганглии: верхний и нижний брыжеечные, чревный узлы).
Нервные узлы содержат ганглионарные нейроны и интернейроны. Передача возбуждения в симпатических ганглиях с преган- глионарного нейрона на нейрон узла осуществляется с помощью медиатора — ацетилхолина, который действует на Н-холиноре- цепторы. В структуре этих рецепторов имеются № + -К + -каналы. Активация Ыа + -К + -каналов вызывает входящий ток ионов натрия и возникновение возбуждающего постсинаптического потенциала (ВПСП). В ганглиях также имеются тормозные адренергические интернейроны — МИФ- клетки. МИФ- клетки (малые интенсивно флуоресцирующие клетки) — небольших размеров нервные клетки, содержащие серотонин.

Рис. 2.2.4. Симпатический отдел вегетативной нервной системы
Ганглионарные нейроны и интернейроны возбуждаются под влиянием преганглионарных нейронов и формируют на ганли- онарных нервных клетках тормозный постсинаптический потенциал (ТПСП). В регуляции передачи возбуждения в симпатических ганглиях помимо этих структур участвуют и другие рецепторы, например М-холинорецепторы, опиатные рецепторы, которые расположены на ганглионарных нейронах.
Постганглионарные волокна образуют немиелиновые аксоны ганглионарных нейронов (тип С, скорость проведения возбуждения около 1 м/с). Потенциал действия (ПД) этих нейронов имеет длительную следовую гиперполяризацию, что свидетельствует о пониженной возбудимости нейрона после спайка потенциала действия. Соотношение числа пре- и постганглионарных волокон составляет 1 : 100. Такое соотношение нервных волокон обеспечивает дивергенцию (расхождение) и мультипликацию (умножение) возбуждения и способствует мобилизации функциональных резервов организма.
Нейроэффекторные синапсы образуются пресинаптическими окончаниями аксона ганлионарных нейронов, имеющими многочисленные варикозные утолщения, каждый из которых образует синапс на эффекторных клетках. Расстояние между ними и эффекторными клетками (синаптическая щель) составляет 10—2000 нм. Типичные постсинаптические мембраны не выражены, рецепторы к медиаторам более равномерно распределены по эффекторной клетке. Медиатор из пресинаптического окончания путем диффузии поступает к многочисленным исполнительным клеткам. В передаче сигнала участвуют симпатические адренергические синапсы и симпатические холинергические синапсы (последние имеются в небольшом количестве).
Симпатические адренергические синапсы содержат основной медиатор норадреналин — 90% (а также адреналин — 7%, дофамин — 3% и комедиатор нейропептид Y). Комедиаторами называют синаптические посредники, характеризующиеся, прежде всего, совместной локализацией, а также совместным высвобождением и общей с медиаторами клеткой-мишенью. Норадреналин действует на а- и p-адренорецепторы, расположенные на постсинаптической мембране. Эффект действия зависит от типа адренорецепторов (например, медиатор через а-рецепторы вызывает сужение кровеносных сосудов, а через pi-рецепторы — их расширение), количества рецепторов и их соотношения на клетке. Действие норадреналина осуществляется через аг и а2- адренорецепторы, каждый из которых имеет несколько подтипов. При стимуляции а-адренорецепторов реакции органов и тканей различаются: в кровеносных сосуды (аь а2) повышается тонус стенки и они сужаются, гладкомышечные клетки кишечника и желудка (ос() расслабляются, гладкомышечные сфинктеры (oci) сокращаются, мочеточники (ось ос2) — сокращаются, а в (3-клетках островков Лангер- ганса поджелудочной железы (ос2) тормозится секреция инсулина.
Эффекты норадреналина, опосредованные oci-рецепторами, реализуются преимущественно через Gq-белки мембраны с последующей активацией фермента фосфолипазы С и других элементов сигнальной системы. Происходит сокращение гладкомышечных клеток сосудов, сфинктеров, радиальной мышцы радужки и других гладкомышечных образований.
Действие норадреналина, опосредованное а2-рецепторами, также начинается преимущественно через Gj-белки мембраны и сопровождается торможением аденилатциклазной системы. В гладкомышечных клетках повышается концентрация Са 2+ и активируются ферменты киназы. Происходит сокращение ГМК, снижение активности ферментов, синтеза гормонов в эндокринных клетках (например, инсулина, ангиотензина II и др.). В жировой ткани тормозятся эффекты ферментов, ускоряющих метаболизм липидов (липаз), в печени уменьшается активность фермента фосфорилазы, одного из главных участников синтеза гликогена (гликогенолиза), а в пресинаптическом окончании нейрона снижается интенсивность синтеза и секреции медиатора норадреналина (саморегуляция синапса).
Действует норадреналин через (3-адренорецепторы (через (3j действие сильное, через |32 — слабое). Локализация и реализуемые через них эффекты: сердце ((3i) — увеличение силы и частоты сокращений; сосуды и бронхи ((32 > pi) — расширение; гладкомышечные клетки кишечника (Р2 > Pi), мочевого пузыря, матки (р2) — расслабление; жировая ткань (|3i) — активируется гидролиз (липолиз); печень ((32) — ферментативный распад гликогена (гликогенолиз) и образование глюкозы из белков (глюконеогенез). Механизмы действия следующие. Норадреналин через p-адренорецепторы и затем мембранный Gs-белок активирует аденилатциклазу, что в итоге приводит к фосфорилирированию различных белков: в сердце тормозится Са 2+ -насос эндоплазматической сети, в результате чего увеличивается концентрация Са 2+ в цитозоле и сила сокращения кардиомиоцита; в ГМК происходят стимуляция Са 2+ -насоса, уменьшение концентрации Са 2+ и расслабление ГМК; фосфорилирование ферментов увеличивает синтез гормонов — либеринов и статинов гипоталамуса, тропных гормонов аденогипофиза, инсулина, фосфорилазы в печени и липазы жировой ткани. В пресинаптическом окончании адренергических синапсов стимулируется синтез и секреция медиатора (саморегуляция синапса).
Инактивация медиатора происходит следующим образом: около 80% норадреналина поступает путем эндоцитоза обратно в преси- наптическое окончание, небольшая часть разрушается моноамино- оксидазой (МАО) пре- и постсинаптической мембран и катехол-Ометилтрансферазой (КОМТ) постсинаптической мембраны, а часть диффундирует в межклеточное пространство и кровь.
Симпатические холинергические синапсы представлены в небольшом количестве (в потовых железах, ГМК сосудов скелетных мышц, мозговом слое надпочечников). Медиатор ацетилхолин действует на М- и Н-холинорецепторы постсинаптической мембраны (активируются также соответственно мускарином и никотином). В синапсах потовых желез и кровеносных сосудах ацетилхолин через М-холинорецепторы вызывает усиление секреции потовых желез и расслабление гладкомышечных клеток сосудов скелетных мышц (расширение сосудов). В мозговом слое надпочечников ацетилхолин действует на Н-холинорецепторы в синапсах на хромаффинных клетках надпочечников, усиливая секрецию катехоламинов. Особенность этой области заключается в том, что нейросекреторные синапсы в надпочечниках образуют преганглионарные симпатические волокна, а хромаффинные эффекторные клетки являются аналогом ганглионарного нейрона. Скопления хромаффинных клеток расположены также на поверхности аорты, в каротидном тельце, в симпатических ганглиях.
В симпатической системе проявляется ауторегуляция передачи сигналов в синапсах. Она обусловлена действием выделяющегося нейромедиатора на пресинаптической мембране того же нервного окончания. Ауторегуляция секреции норадреналина в синапсе осуществляется через рецепторы на пресинаптической мембране (рис. 2.2.5. Механизмы передачи влияний в нервно-эффекторных синапсах вегетативной нервной системыС^). Высокая концентрация норадреналина в синаптической щели и адреналина в крови вызывает через ос2-адренорецепторы торможение высвобождения норадреналина из пресинаптического окончания. Низкая концентрация норадреналина в синаптической щели и адреналина в крови, наоборот, вызывает через (32-адренорецепторы усиление секреции норадреналина из пресинаптического окончания. Через М-холинорецепторы происходит реципрокное торможение ацетилхолином секреции норадреналина при возбуждении парасимпатического отдела.
На основании этих и многих других фактов Л .А. Орбели сформировал представление об адаптационно-трофической функции симпатической нервной системы. Согласно этому представлению симпатические влияния не сопровождаются непосредственно видимым действием, но значительно повышают адаптивные возможности эффектора. Особенно ярко участие симпатической нервной системы обнаруживается в формировании эмоциональных реакций человека, независимо от причин, их вызывающих. Например, радость сопровождается тахикардией, расширением сосудов кожи, а страх — замедлением сердечного ритма, сужением сосудов кожи, повышенным потоотделением, торможением перистальтики кишечника. Гнев вызывает расширение зрачков. Следовательно, в процессе эволюции симпатическая нервная система превратилась в инструмент срочной мобилизации всех ресурсов (интеллектуальных, энергетических и др.) организма как целого в тех случаях, когда возникает угроза самому его существованию.
Возбуждение симпатической нервной системы приводит к изменению гомеостатических констант организма. Это выражается в подъеме кровяного давления, увеличении объема циркулирующей крови за счет ее выхода из депо, поступлении в кровь ферментов, глюкозы, повышении интенсивности метаболизма в органах и тканях, снижении мочеобразования, угнетении функции пищеварительного тракта. Поддержание постоянства этих показателей целиком ложится на парасимпатический и метасимпатический отделы нервной системы. Как видно, объектами управления симпатической нервной системы являются в основном процессы, которые сопровождаются повышенным расходом энергии в организме. Парасимпатический и метасимпатический отделы нервной системы, в отличие от симпатической, способствуют сохранению и накоплению запасов энергии в организме.
Парасимпатический отдел. Центры парасимпатического отдела вегетативной нервной системы расположены в стволе головного мозга и крестцовых сегментах спинного мозга. В стволе мозга расположены парасимпатические ядра III черепного нерва (рефлексы зрачка и хрусталика), верхнее слюноотделительное ядро VII нерва и нижнее слюноотделительное ядро IX нерва (секреция слюнных желез, сосуды слизистой оболочки рта), парасимпатическое ядро X (блуждающего) нерва (органы грудной и брюшной полостей, кроме органов малого таза). В спинном мозге они локализуются в сегментах S2—S4 (органы малого таза и наружные половые органы). В этих центрах расположены преганглионарные нейроны. Область иннервации парасимпатической системы более узкая в сравнении с симпатической нервной системой (например, она не представлена в сосудах кожи) (рис. 2.2.6).

Рис. 2.2.6. Вегетативная парасимпатическая нервная система
Преганглионарные волокна имеют такие же свойства, как и преганглионарные симпатические нейроны (тип В, скорость проведения около 10 м/с). Потенциал действия (ПД) характеризуется длительной следовой деполяризацией, но он, в отличие от симпатических нейронов, имеет большую протяженность.
Нервные узлы (ганглии) парасимпатического отдела нервной системы располагаются вблизи иннервируемого органа, либо непосредственно в его стенке (интрамурально). Механизм передачи возбуждения в ганглиях аналогичен тому, что имеется в симпатических нервных узлах: медиатор ацетилхолин действует на Н-холи- норецептор, содержащий в своем составе № + /К + -канал, открытие которого формирует входящий Na + -TOK и ВПСП.
Постганглионарные волокна (тип С, скорость проведения возбуждения 1 м/с), проводят ПД с длительной следовой гиперполяризацией. В отличие от симпатического отдела соотношение пре- и постганглионарных волокон в вегетативной нервной системе 1 : 1, т.е. нет преобладания дивергенции.
Нейроэффекторные синапсы образованы окончаниями аксонов постганглионарных нейронов на исполнительных клетках органов. В них секретируется медиатор ацетилхолин, который действует на М-холинорецепторы постсинаптической мембраны. М-холи- норецепторы широко представлены в органах и тканях. Активация этих рецепторов в сердце вызывает уменьшение частоты и силы сокращений миокарда, в сосудах приводит к расслаблению их стенки и расширению, а в бронхах, наоборот, сопровождается бронхоспазмом. В желудке, кишечнике, мочевом пузыре М-холинерги- ческие стимулы вызывают сокращение гладкомышечных клеток, а в слюнных, слезных и желудочных железах стимулируют продукцию соответствующих секретов.
Ацетилхолин в проводящей системе сердца реализует свои эффекты через гуанилатциклазную систему. При этом протеинкиназа G активирует К + -каналы. Выходящий из клетки ток ионов калия тормозит медленную диастолическую деполяризацию атипичных кардиомиоцитов, что приводит к уменьшению ритма и проводимости миокарда. В кровеносных сосудах эффект ацетилхолина реализуется через цитозольную гуанилатциклазу. Она активируется образующимся в эндотелии оксидом азота (N0), что приводит к увеличению уровня цГМФ и активации протеинкиназы G. Протеинкиназа G ингибирует ферменты (киназу) легких цепей сократительных элементов (миозина), что приводит к расслаблению ГМК сосудов. В ГМК желудка, кишечника, мочевого пузыря, бронхов и других органов эффект ацетилхолина реализуется через Gq-белок и фосфо- изонитидную сигнальную систему.
Инактивация медиатора: большая часть ацетилхолина разрушается специфическим ферментом — ацетилхолинэстеразой, а оставшийся медиатор диффундирует в межклеточное пространство и кровь.
Ауторегуляция секреции ацетилхолина в синапсе осуществляется через рецепторы на пресинаптической мембране. Высокая концентрация ацетилхолина в синаптической щели вызывает через М-холинорецепторы торможение освобождения ацетилхолина из пресинаптического окончания. Низкая концентрация ацетилхолина в синаптической щели вызывает через Н-холинорецепторы усиление секреции ацетилхолина из пресинаптического окончания. Норадреналин через а2-адренорецепторы вызывает сопряженное (реципрокное) торможение секреции ацетилхолина в синапсе парасимпатического отдела при возбуждении симпатического отдела.
Ведущая роль парасимпатического отдела вегетативной нервной системы заключается в сохранении и восстановлении гомеостаза при его изменениях под влиянием факторов среды, а также регуляции процессов образования сложных органических соединений
(iанаболизма) и запасании пластических и энергетических резервов в организме.
Метасимпатический отдел. Представляет собой морфофункциональный комплекс микроганглиев, расположенных в стенках полых органов, обладающих собственной моторной активностью и соответствующими водителями ритма (сердце, желудочно-кишечный тракт, мочеполовая система).
В зависимости зависит от химической природы медиаторов, типа рецепторов, включаемых внутриклеточных посредников, исходного состояния органов метасимпатический отдел активирует или тормозит их функции. Также как и другие отделы ВНС, метасимпатический отдел оказывает регулирующее влияние на моторику иннервируемых органов, секрецию и всасывание веществ в желудочно-кишечном тракте, местный кровоток, секрецию эндокринных клеток, расположенных в зоне метасимпатической иннервации.
Физиологическое значение метасимпатического отдела ВНС заключается в повышении надежности регуляции висцеральных функций и придании ВНС большей автономности от ЦНС. Мета- симпатическая система разгружает высшие отделы ЦНС от обработки излишней информации, которая необходима для внутриор- ганной регуляции функций. Количество нейронов в ганглиях метасимпатического отдела превышает количество нейронов в спинном мозге.
Читайте также:
