
Медиатор выделяемый окончаниями холинергических нервов
Холинергические механизмы нервной системы - это вещества, которые обеспечивают передачу возбуждения в холинергическом синапсе.
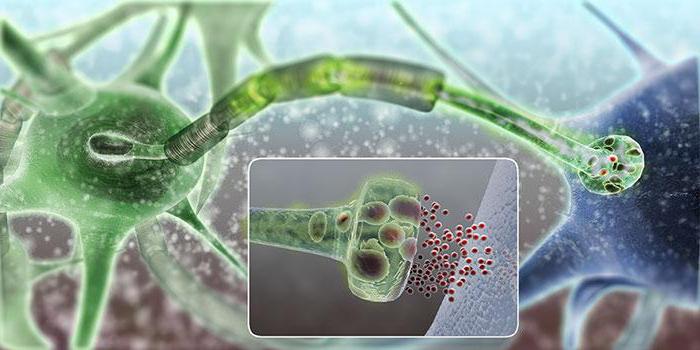
Медиатор ацетилхолин (эфир холина и уксусной кислоты) образуется из аминокислоты холина и ацетил-СоА на пресинаптическом окончании нервноего волокна. Образующийся медиатор поступает в везикулы, а частично может остаться в свободном состоянии. При возбуждении медиатор выделяется из везикул. Процесс выделения медиатора С-зависим. Для нормальной работы синапса необходим запас медиатора, поэтому на пресинаптической мембране идёт ресинтез ацетилхолина. Для этого аминокислота холин выделяется из постсинаптической мембраны, частично из синаптической щели (возврат медиатора). Для образования медиатора необходима энергия метехондрий.
Фермент, способствующий синтезу ацетилхолина - ацетилхолинтрансфераза или холинацетилаза. Этот фермент образуется в теле нейрона и поступает в нервные окончания. Для нормального образования медиатора необходима целостность тела нейрона. Изолированное нервное волокно не может долго выделять медиатор.
Фермент, расщепляющий ацетилхолин - ацетилхолинэстераза. Этот фермент обладает высоким сродстворм к ацетилхолину, который находится в виде комплекса и Х-рецептором. Различают истинную ацетилхолинэстеразу (находится в синапсах и эритроцитах), которая расщепляет ацетилхолин в физиологических концентрациях и ложную ацетилхолинэстеразу (в жидкостях организма - слюне, плазме и т. д.), которая расщепляет ацетилхолин в высоких концентрациях и разрушает еще и различные производные ацетилхолина (курарекодовые препараты). Освобождённый холин с помощью переносчиков поступает на пресимпатическую мембрану, а уксусная кислота и глюкоза поступают в кровь через межтканевую жидкость.
Х-рецепторы - белковые молекулы, обладающие высоким сродством к ацетилхолину.
Существует 2 вида холинорецепторов - М и Н.
М-холинорецепторы - чувствительны к мускалину (яду мухомора) - расположены в основном во внутренних органах, эндокринных железах, сердце, сосудах, дыхательных путях, желудочнокишечном тракте. Они обладают медленным, но продолжительным действием, могут суммировать возбуждение. Существуют 2 вида М-холинорецепторов: одна - во внутренних органах, другая - в эндокринных железах. При возбуждении М-холинорецепторв происходит торможение сердечной деятельности, раширение сосудов, активация деятельности желудочно-кишечного тракта, изменяется секреция некоторых эндокринных желёз.
Н-холинорецепторы - чувствительны к никотину. Располагаются в вегетативных ганглиях, мионевральных синапсах, в хлорофильной ткани надпочечников. Эти рецепторы обладают быстрым, кратковременным действием, не могут суммировать возбуждение. Существует 3 разновидности. За счёт наличия разновидностей рецепторы могут блокироваться различными веществами. В центральной нервной системе больше Н-холинорецепторов. М-холинорецепторы преобладают в области ствола мозга, подкорковых узлах, лимбической системе, ретикулярной формации, гипоталамусе.
Адренергические механизмы нервной системы осуществляются за счет норадреналина - составляет 90 % и других катехоламинов - 10 %.
Предшественник норадреналина - изопропилнораденалин, дофамин. Для синтеза необходимы аминокислоты тиронин, фениламин, которые поступают с постсинапсической мембраны и из тела нейрона. Любые структуры могут образовывать норадреналин, но 95 % его образуется на пресимпатической мембране.
Ферменты синтеза норадреналина - трансаминазы.
Ферменты разрушения ноадреналина - группа катехоламинтрансфераз, часто моноаминоуксусная кислота и моноаминооксидант.
Адренорецепторы - белковые молекулы, обладающие сродством к норадреналину и его производным. Эти рецепторы - наружная субъединица крайней белковой молекулы, внутренняя субъединица может быть ферментом (адемилат- и гуанилатциклазы). При взаимодействии с рецептором изменяется структура молекулы белка и, как следствие, изменяется активность фермента.
Существуют 2 вида адренорецепторов:
Альфа-адренорецепторы - блокируется дегидроэрготамином, обладают повышенной чувствительностью и норадреналину, имеют низкий порог раздражения, при выделении необходимого количества медиатора возбуждаются альфа-рецепторы. Они расположены в некоторых внутренних органах и сосудистой стенке, встречается в центральной нервной системе. Различают альфа 1- и альфа 2-адренорецепторы.
Альфа 1-адренорецепторы - при их возбуждении происходит сужение сосудов, сокращение капсулы селезёнки, матки (особенно беременной), сужение зрачка и т. д. Происходит торможение желудочнокишечного тракта (моторной и секреторной), сокращение сфинктеров.
Альфа 2-адренорецепторы - в основном в центральной нервной системе.
Бетта-адренорецепторы - блокруются бетта-блокаторами (пропранолол). Они обладают высоким порогом раздражения, т. к. имеют меньшее сродство к норадреналину. Чувствительны к различным производным норадреналина (изопротеренолол).
Бетта 1-адренорецепторы - в миокарде; при их возбуждении увеличивается сила сердечных сокращений, ускоряются обменные процессы в миокарде, несколько увеличивается частота сердечных сокращений.
Бетта 2-адренорецепторы - в сосудах, внутренних органах, эндокринных железах. При их возбуждении обеспечивается тормозной эффект, расширение сосудов (коронарных, скелетных мышц), расслабление гладких мышц, дыхательных путей. В сосудах могут встречаться альфа 1- и бетта 2-рецепторы.
Альфа 1-рецепторы обеспечивают сужение, а бетта 2 - расширение сосудов. Эффект зависит от: количества медиаторов, количества рецепторов данного вида.
Симпатические и парасимпатические нервные волокна секретируют в основном один из двух синаптических медиаторов — ацетилхолин или норадреналин. Волокна, секретирующие ацетилхолин, называют холинергическими, волокна, секретирующие норадреналин, называют адренергическими (термин, происходящий от адреналина, — альтернатива эпинефрину).
Все преганглионарные нейроны (и симпатической, и парасимпатической нервных систем) являются холинергическими. Ацетилхолин или подобные ему вещества при действии их на ганглии возбуждают симпатические и парасимпатические постганглионарные нейроны. Все или почти все постганглионарные нейроны парасимпатической системы — также холинергические. С другой стороны, большинство постганглионарных симпатических нейронов являются адренергическими.
Однако постганглионарные симпатические нервные волокна, идущие к потовым железам, мышцам, поднимающим волосы, и к очень небольшому числу кровеносных сосудов, являются холинергическими.
Таким образом, нервные окончания парасимпатической системы все или практически все секретируют ацетилхолин. Почти все симпатические терминали секретируют норадреналин, но немногие из них секретируют ацетилхолин. Эти вещества при введении их в кровь, в свою очередь, действуют на разные органы, вызывая соответствующие парасимпатические или симпатические эффекты, поэтому ацетилхолин называют парасимпатическим медиатором, а норадреналин — симпатическим медиатором.

Секреция ацетилхолина и норадреналина постганглионарными нервными окончаниями. Некоторые постганглионарные вегетативные нервные окончания, особенно парасимпатические, подобны таковым в нервно-мышечном соединении, но гораздо мельче. Однако многие парасимпатические волокна и почти все симпатические волокна лишь слегка касаются эффекторных клеток, проходя мимо них в иннервируемых органах; в некоторых случаях они заканчиваются в соединительной ткани, прилежащей к стимулируемым клеткам.
Там, где эти нервные терминали проходят над стимулируемыми клетками или рядом с ними, они обычно имеют варикозные расширения, называемые варикозами; именно в этих варикозах синтезируются и хранятся в везикулах медиаторы (ацетилхолин или норадреналин). Кроме того, в варикозах имеется большое количество митохондрий, поставляющих аденозинтрифосфат, необходимый для обеспечения энергией синтеза ацетилхолина и норадреналина.
Когда потенциал распространяется вдоль нервных терминалей, процесс деполяризации увеличивает проницаемость мембраны волокна к ионам кальция, позволяя им диффундировать в терминаль, или варикозы, опорожняя их содержимое наружу. Так секретируется медиатор.
Синтез ацетилхолина, его разрушение после секреции и длительность его действия. Ацетилхолин синтезируется в терминальных окончаниях и варикозах холинергических нервных волокон, где он хранится в везикулах в высококонцентрированной форме, пока не выделится.
После выделения ацетилхолина из холинергического нервного окончания в ткань он сохраняется в ткани в течение нескольких секунд, выполняя свою функцию передачи нервного сигнала. Затем он расщепляется на ион уксусной кислоты (ацетат) и холин под действием фермента ацетилхолинестеразы, который связан с коллагеном и глюкозамингликанами местной соединительной ткани. Этот механизм передачи сигнала ацетилхолином и последующего его разрушения аналогичен процессам, которые происходят в терминалях соматических нервных волокон в нервно-мышечных соединениях.
Образовавшийся холин затем транспортируется назад в окончание нервной терминали, где он снова используется для синтеза новых молекул ацетилхолина.
Медиатор ацетилхолин - сложный эфир четвертичного аминоспирта холина и уксусной кислоты. Синтезируется в цитоплазме окончаний холинергических нервов при участии холинацетилазы и депонируется в везикулах. Изменение потенциала мембраны нервного окончания, вызываемого импульсом, приводит к поступлению в цитоплазму ионов кальция, которые, активируя кальмодулин, вызывают высвобождение медиатора из везикулы и выброс его в синаптическую щель. Ацетилхолин связывается с холинорецепторами постсинаптической мембраны и гидролизуется (разрушается) внутрисинаптической ацетилхолинэстеразой (истинной холинэстеразой). В плазме крови имеется другая холинэстераза, называемая ложной или псевдохолинэстеразой (бутирилхолинэстераза). Последняя образуется в печени и, поступая в кровь, легко разрушает соединения, представляющие собой низкомолекулярные сложные эфиры, в том числе ацетилхолин и некоторые лекарства, например, новокаин, дикаин, дитилин, атропин и др.
Взаимодействие ацетилхолина с холинорецепторами изменяет проницаемость постсинаптических мембран для ионов. Ионы натрия входят, а калия выходят из клетки, мембранный потенциал снижается (деполяризация), клетка возбуждается; в нейроне возбуждение в виде потенциала действия распространяется по аксону. Почти одновременно ацетилхолин разрушается, и рецепторы освобождаются, полярность мембраны восстанавливается (реполяризация), нормализуется содержание натрия и калия - за счет работы натрий-калиевого насоса избыток натрия выходит из клетки в обмен на калий.
Холинорецепторы неоднородны, они проявляют неодинаковую чувствительность к различным веществам. Выделяют холинорецепторы, реагирующие на мускарин (яд гриба мухомора) - их называют М-холинорецепторы (мускариночувствительные), и Н- холинорецепторы (никотиночувствительные), они возбуждаются малыми дозами никотина.
М-холинорецепторы локализованы в мембранах:
1) клеток, иннервируемых постганглионарными парасимпатическими волокнами (проводящая система сердца, глаз, железы внешней секреции, гладкомышечные клетки, в том числе бронхов и желудочно-кишечного тракта);
2) клеток потовых желез, иннервируемых постганглионарными симпатическими волокнами холинергического типа;
3) нейронов некоторых отделов центральной нервной системы (кора головного мозга, ретикулярная формация и др.).
1) в нейронах симпатических и парасимпатических ганглиев;
2) в синокаротидных клубочках (расположены в месте деления сонных артерий);
3) в хромаффинных клетках мозгового слоя надпочечников;
4) в клетках скелетных мышц;
5) в нейронах некоторых отделов ЦНС. Лекарственные вещества, действующие подобно ацетилхолину, называют холиномиметиками (от греческого mimeticos -подражающий) и подразделяют на:
1) М- и Н-холиномиметики (возбуждающие и М-, и Н-холинорецепторы);
2) М-холиномиметики (возбуждающие М-холинорецепторы);
3) Н-холиномиметики (возбуждающие Н-холинорецепторы); Лекарственные средства, блокирующие холинорецепторы - холиноблокаторы, или холинолитики (от греч. lyticos -разрушающий) включают:
1) М- и Н-холинолитики - блокирующие М- и Н-холинорецепторы;
2) М-холинолитики - блокирующие М-холинорецепторы;
3) Н-холинолитики - блокирующие Н-холинорецепторы.
Большинство холинергических средств имеет общие с ацетилхолином особенности химической структуры - именно поэтому связываются с холинорецептором. Они являются основаниями, эфирами и содержат третичные или четвертичные атомы азота. Соединения третичного азота не диссоциируют, хорошо растворимы в жирах. легко всасываются в желудочно-кишечном тракте, проникают через гематоэнцефалический барьер и поэтому могут оказывать действие на центральную нервную систему. Четвертичные азотсодержащие соединения имеют четырехвалентный азот, у которого три валентности прочно связаны, а четвертая может образовывать ионную связь с анионами, например, кислот. Эти соединения плохо растворимы в жирах, практически не всасываются в пищеварительном тракте, гематоэнцефалический барьер не проходят, а значит не влияют на головной и спинной мозг. Для них характерны, в основном, периферические эффекты.
- 2.1.2.1.1. Холиномиметические средства
- 2.1.2.1.1.1. Средства, стимулирующие М-холинорецепторы (М-холиномиметики)
- 2.1.2.1.1.2. Средства, стимулирующие Н-холинорецепторы (Н-холиномиметики)
- 2.1.2.1.1.3. Средства, стимулирующие М- и Н-холинорецепторы (М-, Н-холиномиметики)
- 2.1.2.1.2. Средства, блокирующие М- и Н-холинорецепторы
- 2.1.2.1.2.1. Средства, блокирующие М-холинорецепторы (М-холинолитики)
- 2.1.2.1.2.2. Средства, блокирующие Н-холинорецепторы (Н-холинолитики)
- 2.1.2.1.2.2.1. Ганглиоблокаторы
- 2.1.2.1.2.2.2. Курареподобные средства (миорелаксанты периферического действия)
| Листать назад | Оглавление | Листать вперед |
- Лекарства и субстанции
- Указатель лекарств и субстанций
- Указатель действующих веществ
- Производители
- Фармакологические группы
- Классификация фармакологических групп
- Указатель фармакологических групп
- АТХ классификация
- Классификация лекарственных форм
- Справочник болезней
- Международная классификация болезней (МКБ-10)
- Указатель болезней и состояний
- Взаимодействие лекарств (действующих веществ)
- Указатель фармакологических действий
- Проверка подлинности упаковок по 3D
- Поиск регистрационных удостоверений
- БАДы и другие ТАА
- БАДы
- Указатель БАДов
- Классификация БАДов
- Другие ТАА
- Указатель других ТАА
- Классификация других ТАА
- БАДы
- Цены
- Цены на ЖНВЛП
- Цены на лекарства и другие ТАА в Москве
- Цены на лекарства и другие ТАА в Санкт-Петербурге
- Цены на лекарства и другие ТАА в регионах
- Новости и события
- Новости
- События
- Пресс-релизы фармкомпаний
- Архив мероприятий
- Продукты и сервисы
- Цены на ЖНВЛП
- 3D-упаковки
- Гармонизация
- Забраковка
- Взаимодействие
- Фармэквивалентность
- Электронные версии справочников для врачей
- Мобильные приложения
- Поиск лечебных учреждений в РФ
- Библиотека
- Книги
- Cтатьи
- Нормативные акты
- О компании
- Аптечка
- Интернет-магазин
Все права защищены. Не разрешается коммерческое использование материалов. Информация предназначена для медицинских специалистов.
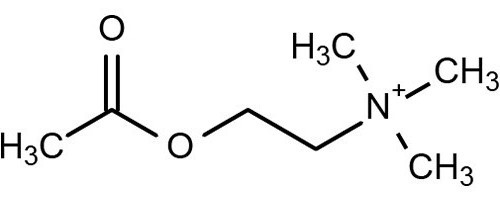
Ацетилхолин – это передатчик нервного возбуждения в ЦНС, окончаниях парасимпатических нервов и вегетативных ганглиях. Он выполняет важнейшие задачи в процессах жизнедеятельности. Аналогичными функциями обладают аминокислоты, гистамин, дофамин, серотонин, адреналин. Ацетилхолин считается одним из важнейших передатчиков импульсов в мозг. Рассмотрим это вещество подробнее.
Общие сведения
Окончания волокон, от которых медиатор ацетилхолин осуществляет передачу, именуются холинергическими. Кроме этого, существуют специальные элементы, с которыми он взаимодействует. Они называются холинорецепторами. Эти элементы представляют собой сложные молекулы белка – нуклеопротеиды. Рецепторы ацетилхолина отличаются тетрамерной структурой. Они локализуются на внешней поверхности плазматической (постсинаптической) мембраны. По своей природе эти молекулы неоднородны.
В экспериментальных исследованиях и в медицинских целях используется препарат "Ацетилхолин-хлорид", представленный в растворе для инъекций. Другие лекарственные средства на основе этого вещества не выпускаются. Существуют синонимы препарата: "Миохол", "Ацеколин", "Цитохолин".
Классификация холиновых белков
Некоторые молекулы находятся в районе холинергических постганглионарных нервов. Это область гладкой мускулатуры, сердца, желез. Они называются м-холинорецепторами – мускариночувствительными. Другие белки расположены в районе ганглионарных синапсов и в нервно-мышечных соматических структурах. Они именуются н-холинорецепторами – никотиночувствительными.
Пояснения
Приведенная выше классификация обуславливается спецификой реакций, которые возникают, когда взаимодействуют эти биохимические системы и ацетилхолин. Это , в свою очередь, объясняет причины некоторых процессов. Например, снижение давления, усиленную секрецию желудочных, слюнных и прочих желез, брадикардию, сужение зрачков и пр. при влиянии на мускариночувствительные белки и сокращение скелетных мышц и пр. при воздействии на никотиночувствительные молекулы. При этом в последнее время ученые начали разделять м-холинорецепторы на подгруппы. Наиболее изучена сегодня роль и локализация м1- и м2-молекул.
Специфика влияния
Ацетилхолин – это не избирательный элемент системы. В той или иной степени он воздействует и на м-, и на н-молекулы. Интерес представляет мускариноподобное влияние, которое оказывает ацетилхолин. Это воздействие проявляется в замедлении сердечного ритма, расширении кровеносных сосудов (периферических), активизации перистальтики кишечника и желудка, сокращении мышц матки, бронхов, мочевого, желчного пузыря, интенсификации секреции бронхиальных, потовых, пищеварительных желез, миозе.
Сужение зрачка
Круговая мышца радужной оболочки, иннервируемая постганглионарными волокнами в глазодвигательном нерве, начинает усиленно сокращаться одновременно с ресничной. При этом имеет место расслабление цинновой связки. В результате возникает спазм аккомодации. Сужение зрачка, связанное с влиянием ацетилхолина, как правило, сопровождается понижением внутриглазного давления. Данный эффект частично обуславливается расширением оболочки в шлеммовом канале и фонтановых пространств на фоне миоза и уплощения радужной оболочки. Это способствует улучшению оттока жидкости из внутренних глазных сред.
Благодаря возможности понижать внутриглазное давление, как ацетилхолин, препараты на основе других подобных ему веществ используются при лечении глаукомы. К ним, в частности, относят антихолинэстеразные средства, холиномиметики.
Никотиночувствительные белки
Никотиноподобное действие ацетилхолина обуславливается его участием в процессе передачи сигналов с преганглионарных нервных волокон на постганглионарные, находящиеся в вегетативных узлах, и с двигательных окончаний на поперечнополосатые мышцы. В малых дозах вещество выступает в качестве физиологического передатчика возбуждения. Если ацетилхолин повышен , то может развиться стойкая деполяризация в районе синапсов. Также существует вероятность блокирования передачи возбуждения.
Ацетилхолин в организме играет роль передатчика сигналов в различных мозговых отделах. В малой концентрации он может облегчать, а в большой – замедлять синаптическую трансляцию импульсов. Изменения обмена вещества могут способствовать развитию мозговых нарушений. Антагонисты, которым противопоставляется ацетилхолин, - препараты психотропной группы. При их передозировке может возникнуть нарушение высших нервных функций (галлюциногенный эффект и пр.).
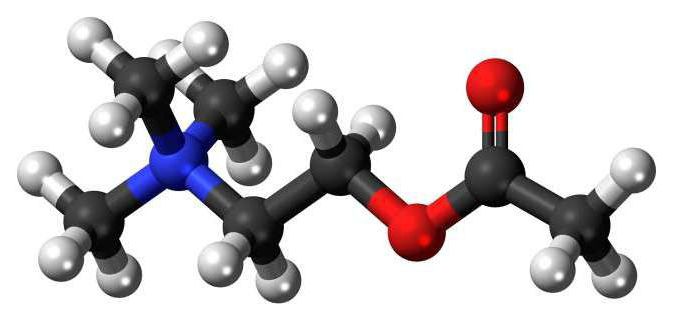
Синтез ацетилхолина
Он происходит в цитоплазме в нервных окончаниях. Запасы вещества располагаются в пресинаптических терминалях в виде пузырьков. Возникновение потенциала действия приводит к высвобождению ацетилхолина из нескольких сотен "капсул" в синаптическую щель. Вещество, выделяющееся из пузырьков, связывается на постсинаптической мембране со специфическими молекулами. Это повышает ее проницаемость для натриевых, кальциевых и калиевых ионов. В результате возникает возбуждающий постсинаптический потенциал. Влияние ацетилхолина ограничивается посредством его гидролиза с участием фермента ацетилхолиэстеразы.
Физиология никотиновых молекул
Первому описанию способствовал внутриклеточный отвод электрических потенциалов. Никотиновый рецептор стал одним из первых, на который удалось записать токи, пропускаемые через единичный канал. В открытом состоянии сквозь него могут проходить ионы К+ и Na+, в меньшей степени двухвалентные катионы. При этом проводимость канала выражена в постоянной величине. Продолжительность открытого состояния, тем не менее, выступает характеристикой, зависящей от напряжения потенциала, приложенного к рецептору. При этом последний стабилизируется при переходе от деполяризации мембраны к гиперполяризации. Кроме этого, отмечается явление десенсетизации. Оно возникает при продолжительной аппликации ацетилхолина и прочих антагонистов, снижающей чувствительность рецептора и увеличивающей длительность открытого состояния канала.
Электрическое раздражение
Дигидро-β-эритроидин блокирует никотиновые рецепторы головного мозга и нервных ганглий при проявлении ими холинергического ответа. Для них также характерно высокоафинное сродство с тритий-меченным никотином. Чувствительные нейронные рецепторы αBGT в гиппокампе отличаются низкой восприимчивостью ацетилхолина, в отличие от нечувствительных αBGT-элементов. Оборотным и селективным конкурентным антагонистом первых выступает метилликаконитин.
Отдельные производные анабезиина провоцируют селективное активационное воздействие на группу αBGT-рецепторов. Проводимость их ионного канала достаточно высока. Эти рецепторы отличаются уникальными вольт-зависимыми характеристиками. Общеклеточный ток при участии деполяризационных величин эл. потенциала указывает на уменьшение пропуска ионов через каналы.
Данное явление при этом регулируется содержанием в растворе элементов Mg2+. Этим данная группа отличается от рецепторов мышечных клеток. Последние не претерпевают каких-либо изменений тока ионов при корректировке величин мембранного потенциала. При этом а N-метил-D-аспартатный рецептор, обладающий относительной проницаемостью для элементов Са2+, показывает обратную картину. При увеличении потенциала до гиперполяризующих значений и повышении содержания ионов Mg2+ ионный ток блокируется.
Особенности мускариновых молекул
М-холинорецепторы относятся к классу серпентивных. Они передают импульсы через гетеротримерные G-протеины. Группа мускариновых рецепторов была выявлена благодаря их свойству связывать алкалоид мускарин. Опосредованно эти молекулы были описаны в начале 20-го столетия при изучении эффектов кураре. Непосредственное исследование этой группы началось в 20-30 гг. того же века после идентификации соединения ацетилхолина как нейромедиатора, поставляющего импульс в нервно-мышечные синапсы. М-белки активизируются под влиянием мускарина и блокируются атропином, н-молекулы активируются под воздействием никотина и блокируются кураре.
Спустя время в обеих группах рецепторов было выявлено большое количество подтипов. В нервно-мышечных синапсах присутствуют только никотиновые молекулы. Мускариновые рецепторы обнаруживаются в клетках желез и мускулатуры, а также - вместе с н-холинорецепторами - в нейронах ЦНС и нервных ганглиях.

Функции
Мускариновые рецепторы обладают целым комплексом различных свойств. В первую очередь они располагаются в автономных ганглиях и отходящих от них постганглиозных волокнах, направленных к органам-мишеням. Это указывает на участие рецепторов в трансляции и модуляции парасимпатических эффектов. К ним, например, относят сокращение гладких мышц, расширение сосудов, усиление секреции желез, снижение частоты сокращений сердца. Холинергические волокна ЦНС, в составе которых присутствуют интернейроны и мускариновые синапсы, сконцентрированы преимущественно в коре мозга, гиппокампе, ядрах ствола, стриатуме. В других участках они обнаруживаются в меньшем количестве. Центральные м-холинорецепторы влияют на регуляцию сна, памяти, обучения, внимания.
Эфферентные нервы и синапсы делятся на холинергические и адренергические, в зависимости от синтезирующегося в окончаниях и выделяющегося в синаптическую щель медиатора.
Холинергическими являются все двигательные, все преганглионарные (парасимпатические и симпатические), постганглионарные парасимпатические волокна, а также постганглионарные симпатические нервные волокна, иннервирующие потовые железы.
Среди холинорецепторов выделяют мускаринчувствительные (м-холинорецепторы) и никотинчувствительные (н-холинорецепторы)– по названиям естественныхалкалоидов, которые оказывают избирательное действие на соответствующие холинорецепторы. Мускариновые холинорецепторы, в свою очередь, могут быть м1-, м2- и м3-типа в зависимости от того, в каких органах или тканях они преобладают.
К адренергическим относятся только постганглионарные симпатические нервные волокна (исключая иннервацию потовых желез). Средства, влияющие на передачу импульса в холинергических синапсах, называют холинергическими, в адренергических - адренергическими.
Адренорецепторы, исходя из различной чувствительности их к химическим соединениям, подразделяют на альфа- и бета-адренорецепторы, которые тоже в зависимости от локализации имеют несколько разновидностей. (альфа1-, альфа2-; бета1-, бета2-)
*Ацетилхолин, переключает возбуждение с преганглионарных волокон на постганглионарные как в симпатической так и в парасимпатической системах (в ПСНС также АЦХ учавствует и в передаче возбуждения на орган). Чувствительными к АЦХ являются м- и н-холинорецепторы. Норадреналин взаимодействует с адренорецепторами. Результат – возбуждение симпатического волокна, зависящее от популяции адренорецепторов; преобладает в органе на постсинаптической мембране. Чувствительными к НА являются альфа- и бета-адренорецепторы.
Некоторые препараты, улучшающие активность мозга (фенилпирацетам) строятся на увеличении содержания медиаторов, что благобриятно действует на консолидацию памяти, концентрацию внимания, повышает скорость передачи информации по нервному волокну, благодаря НА функционируют центры удовольствия.
Холинореактивные системы делят на две группы: М-холинорецепторы (М-холинореактивные системы), характеризующиеся чувствительностью к мускарину, и Н-холинорецепторы (холинореактивные системы), более чувствительные к никотину. М-холинорецепторы расположены в органах с гладкой мускулатурой, в сердечной мышце, секреторных железах (органы с постганглионарной иннервацией) и в центральной нервной системе. Н-хслинорецепторы (на которых оказывают возбуждающее действие малые дозы никотина) находятся в вегетативных ганглиях, хромаффинных клетках надпочечников (медуллярная часть), в поперечнополосатой мускулатуре, в центральной нервной системе.
М-холинореактивные системы значительно более чувствительны к блокирующему действию атропина и скополамина, чем Н-холинореактивные. Однако есть данные, что Н-холинореактивные системы в ганглиях и поперечнополосатой мускулатуре различны. В то время как в ганглиях Н-холинореактивные системы блокируются некоторыми метониевыми соединениями (так называемые ганглиоблокаторы), в поперечнополосатой мускулатуре они избирательно блокируются препаратами кураре и некоторыми курареподобными ядами.
Ацетилхолин играет важную роль в функциональном состоянии центральной нервной системы. При раздражении периферических нервов или некоторых областей коры больших полушарий выделяется ацетилхолин в свободной активной форме (Burgen, Mcintosh, М. Я. Михельсон и др.). Как электрофизиологические исследования, так и исследования поведенческих реакций показывают, что повышенное содержание ацетилхолина в центральной нервной системе, вызываемое физиологическим раздражителем или введением ингибитора фермента, разрушающего ацетилхолин, связано с явлениями возбуждения. Локальное нанесение ацетилхолина на кору больших полушарий вызывает разряды. Под влиянием антихолинэстеразных веществ у животных быстрее вырабатываются рефлексы. Elliott считает, что определенное количество ацетилхолина необходимо для нормальной деятельности центральной нервной системы. Скопление большого количества ацетилхолина вызывает судороги приступообразного характера, обусловленные в большой степени гиперактивацией деятельности центральной нервной системы. Впрочем, данные, касающиеся этого вопроса, противоречивы.
Химическая передача возбуждения в ганглиях симпатической нервной системы. Ацетинхолин как передатчик возбуждения в ганглиях. Роль холиностеразы. Ганглиоблокирующие вещества и их роль в лекарственной терапии.
В ганглиях симпатической НС паравертебральный ганглий располагается по обе стороны от позвоночника. Здесь располагаются синапсы, и возбуждение переключается с преганглионарных нейронов на постганглионарные нейроны с помощью АЦХ и холинорецепторов.
В ганглиях парасимпатической НС преганглионарные волокна несут возбуждение к органу, на органе находится синапс, где переключается возбуждение с преганглионарного волокна на постганглионарный нейрон. От нейрона короткое постганглионарное волокно. В синапсах ганглия медиатор АЦХ. М- и н-холинорецепторы на постсинаптической мембране распознают АЦХ и передают возбуждение. *Холинэстераза в холинэргических синапсах катализирует гидролиз ацетилхолина, и, как следствие, прекращает влияние данного медиатора на холинорецептор.
* Под действием ганглиоблокаторов вегетативные нервные узлы становятся малочувствительными не только к АЦХ, но и к различным холинергическим раздражителям (никотин, мускарин, лобелин, цитизин и др.). Блокируются симпатические (учавствуют адренолитики: бета1-бутанол, альфа1-празоин и др.) и парасимпатические (учавствуют атропин, скопалин) узлы, однако для разных препаратов в разной последовательности и с различной силой. Первоначально ганглиоблокаторы относительно широко применяли для лечения гипертонической болезни, язвенной болезни желудка и двенадцатиперстной кишки, других заболеваний. Сегодня их иногда используют для купирования гипертонических кризов и в комплексной терапии язвенной болезни.
Читайте также:
