
Возбуждение и торможение в цнс впсп и тпсп
В возбуждающих синапсах нервной системы медиатором может являться ацетилхолин, норадреналин, дофамин, серотонин, глугаминовая кислота, вещество Р, а также большая группа других веществ, являющихся, если не медиаторами в прямом значении, то во всяком случае модуляторами (меняющими эффектиьность) синаптической передачи. Возбуждающие медиаторы вызывают появление на постсинаптической мембране возбуждающего постсинаптического потенциала (ВПСП). Его формирование обусловлено тем, что медиатор-рецепторный комплекс активирует Na- каналы мембраны (а также вероятно и Са-каналы) и вызывает за счет поступления натрия внутрь клетки деполяризацию мембраны. Одновременно происходит и уменьшение выхода из клетки ионов К+Амплитуда одиночного ВПСП однако довольно мала, и для уменьшения заряда мембраны до критического уровня деполяризации необходима одновременная активация нескольких возбуждающих синапсов. ВПСП, образуемые на постсинаптической мембране этих синапсов, способны суммироваться, т.е. усиливать друг друга, приводя к росту амплитуды ВПСП (пространственная суммация). Растет амплитуда ВПСП и при увеличении частоты поступающих к синапсу нервных импульсов (временная суммация), что повышает число выводимых в синаптическую щель квантов медиатора. Процесс спонтанной регенеративной деполяризации возникает в нейроне обычно в месте отхождения от тела клетки аксона, в так называемом аксонном холмике, где аксон еше не покрыт миелином и порог возбуждения наиболее низкий. Таким образом, ВПСП, возникающие в разных участках мембраны нейрона и на его дендритах, распространяются к аксонному холмику, где суммируются, деполяризуя мембрану до критического уровня и приводя к появлению потенциала действия. Тормозной постсинаптический потенциал (ТПСП)В тормозных синапсах обычно действуют другие, тормозные, медиаторы. Среди них хорошо изученными являются аминокислота глицин (тормозные синапсы спинного мозга), гамма-аминомасляная кислота (ГАМК) — тормозной медиатор в нейронах головного мозга. Вместе с тем, тормозной синапс может иметь тот же медиатор, что и возбуждающий, но иную природу рецепторов постсинаптической мембраны. Так, для ацетилхолина, биогенных аминов и аминокислот на постсинаптической мембране разных синапсов могут существовать как минимум два типа рецепторов, и, следовательно, разные медиатор-рецепторные комплексы способны вызывать различную реакцию хемочувствительных рецепторуправляемых каналов. Для тормозного эффекта такой реакцией может являться активация калиевых каналов, что вызывает увеличение выхода ионов калия наружу и гиперполяризацию мембраны. Аналогичный эффект во многих тормозных синапсах имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. Возникающий при гиперполяризации сдвиг мембранного потенциала получил название тормозного постсинаптического потенциала (ТПСП). На рис.3.5 показаны отличительные черты ВПСП и ТПСП. Увеличение частоты нервных импульсов, приходящих к тормозному синапсу, также как и в возбуждающих синапсах, вызывает нарастание числа квантов тормозного медиатора, выделяющихся в синаптическую щель, что, соответственно, повышает амплитуду гиперполяризационного ТПСП. Вместе с тем, ТПСП не способен распространяться по мембране и существует только локально. В результате ТПСП уровень мембранного потенциала удаляется от критического уровня деполяризации и возбуждение становится либо вообще невозможным, либо для возбуждения требуется суммация значительно больших по амплитуде ВПСП, т.е. наличие значительно больших возбуждающих токов. При одновременной активации возбуждаюших и тормозных синапсов резко падает амплитуда ВПСП, так как деполяризующий поток ионов Na+компенсируется одновременным выходом ионов К+ в одних видах тормозных синапсов или входом ионов СГ в других, что называют шунтированием ВПСП.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.
ВПСП – возбуждающим постсинаптическим потенциалом
ТПСП – тормозным постсинаптическим потенциалом
ГАМК – гамма-аминомасляной кислоты
Си́напс— место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём посредством прохождения ионов из одной клетки в другую.
Медиатор, находящийся в пузырьках, выделяется в синаптическую щель с помощью экзоцитоза. Его выделение происходит небольшими порциями – квантами. Небольшое количество квантов выходит из окончания и в состоянии покоя. Когда нервный импульс, т.е. ПД, достигает пресинаптического окончания, происходит деполяризация его пресинаптической мембраны. Открываются ее кальциевые каналы и ионы кальция входят в синаптическую бляшку. Начинается выделение большого количества квантов нейромедиатора. Молекулы медиатора диффундируют через синаптическую щель к постсинаптической мембране и взаимодействуют с её хеморецепторами. В результате образования комплексов медиатор-рецептор, в субсинаптической мембране начинается синтез так называемых вторичных посредников (в частности, цАМФ). Эти посредники активируют ионные каналы постсинаптической мембраны. Поэтому такие каналы называют хемозависимыми или рецепторуправляемыми. Т.е. они открываются при действии ФАВ на хеморецепторы. В результате открывания каналов изменяется потенциал субсинаптической мембраны. Такое изменение называется постсинаптическим потенциалом.
В ЦНС возбуждающими являются холин-, адрен-, дофамин-, серотонинергические синапсы и некоторые другие. При взаимодействии их медиаторов с соответствующими рецепторами, открываются хемозависимые натриевые каналы. Ионы натрия входят в клетку через субсинаптическую мембрану. Происходит ее местная или распространяющаяся деполяризация. Эта деполяризация называется возбуждающим постсинаптическим потенциалом (ВПСП).
Тормозными являются глицин- и ГАМК-ергические синапсы. При связывании медиатора с хеморецепторами, активируются калиевые или хлорные хемозависимые каналы. В результате ионы калия выходят из клетки через мембрану.
Ионы хлора входят через нее. Возникает только местная гиперполяризация субсинаптической мембраны. Она называется тормозным постсинаптическим потенциалом (ТПСП).
Величина ВПСП и ТПСП определяется количеством квантов медиатора, выделившихся из терминали, а, следовательно, частотой нервных импульсов. Т.е. синаптическая передача не подчиняется закону "все или ничего". Если количество выделившегося возбуждающего медиатора достаточно велико, то в субсинаптической мембране может генерироваться распространяющийся ПД. ТПСП, независимо от количества медиатора не распространяется за пределы субсинаптической мембраны.
В24. Фундаментальные принципы работы мозга. Свойства нервных центров: одностороннее поведение возбуждения, суммация, трансформация ритма, последействие, утомляемость, окклюзия, облегчение.
Мозг – главнее, потому что:
-- он мощнее и разнообразнее, чем осознаваемая человеком часть мыслительного процесса,
-- он принимает решение самостоятельно без участия сознания, и мы не всегда понимаем, как он это делает,
Принципы работы мозга
Особенность работы мозга человека такова, что о норме в рамках этой темы следует говорить с большой осторожностью. Грань между гениальностью и патологией настолько тонка, что почти незаметна. Психические и нервные расстройства фиксируются уже настолько часто, что начали опережать по количеству сердечно-сосудистые заболевания и онкологию. Тем не менее, существуют нормативные показатели для работы мозговых волн, различные отклонения в регистрации которых дают возможность установить патологии развития.
Мозговые волны
Альфа-уровень работы мозга с частотой 8-13 Гц у 95% здоровых людей регистрируется в состоянии расслабленного бодрствования главным образом в областях затылка и темени.
Бета-ритм. Частота работы мозга 14-40 Гц. В норме имеет слабовыраженные колебания с амплитудой до 3-7 мкВ в областях передних и центральных извилин. Возникает при бодрствовании во время наблюдения или при концентрации на решении проблем.
Гамма-волна возникает при решении задач, требующих максимальной сосредоточенности. Колебания от 30-100 Гц в теменной, височной, фронтальной и прецентральной областях.
Дельта-ритм с колебаниями 1-4 Гц связан с медленными восстановительными процессами и низкой активностью.
Тета-ритм. Его частота – 4-8 Гц с регистрацией в гиппокампе и фронтальных зонах. Возникает при переходе расслабленного бодрствования в сонливость.
Принцип рефлекторной работы
Рефлекс – это реакция организма на раздражение рецепторов (чувствительных образований), выполнение которой происходит с участием нервной системы.
Рене Декартом в 17 веке был открыт рефлекторный принцип нервной деятельности в целом. А предположение о рефлекторной деятельности высших отделов мозга, то есть, принцип рефлекторной работы мозга был открыт И. Сеченовым уже в 19 веке. И. Павлов разработал пути экспериментального объективного исследования функций коры и методику выработки условных рефлексов на безусловные. Развивая эти представления, П. Анохин создал концепцию функциональной системы, в рамках которой утверждается, что в каждый момент времени складывается сложная система – временное объединение чувствительных рецепторов, нервных элементов структур головного мозга с исполнительными органами.
Свойство нервных центров: одностороннее проведение возбуждения, суммация, трансформация ритма, последействие, утомляемость, окклюзия, облегчение.
Дата добавления: 2018-11-24 ; просмотров: 1693 ;
Тормозные синапсы содержат медиатор, вызывающий торможение нейрона путем генерации ТГ1СП. Эго приводит к уменьшению величины ВПСП и снижению либо предотвращению возбуждения нейрона. Тормозные синапсы могут быть образованы как на мембране самого нейрона, так и на мембране аксонов, подходящих к нейрону. Первый тип синапсов осуществляет постсинаптическое торможение, а второй — пресинаптическое.
Тормозные ГАМКергические синапсы, как правило, располагаются на соме нейронов ближе к аксонному холмику, в отличие от возбуждающих синапсов, сосредоточенных главным образом на дендритах. Например, при обычном для ряда центральных нейронов потенциале покоя около -70 мВ ТПСП имеет вид небольшой гиперполяризации с амплитудой 1—5 мВ. Однако в большинстве клеток распределение анионов хлора по обе стороны мембраны таково, что его равновесный потенциал совпадает с мембранным потенциалом клетки, а в некоторых нейронах значение Ес| оказывается ниже величины мембранного потенциала. Поэтому при активации и открытии под действием ГАМК хлорных каналов ГАМК- рецепторов трансмембранный хлорный ток может не возникать или за счет выхода анионов хлора наружу приводить к развитию деполяризующего ГАМКергического ТПСП. Существуют три варианта постсинаптического торможения (рис. 3.33).

Рис. 3.33. Постсинаптическое торможение:
приведены примеры генерации ГАМКергических ТПСП (гиперполяризационных, деполяризационных и равных нулю), вызванных хлорным током. Направленность ТПСП зависит от соотношения потенциала на мембране и равновесного хлорного потенциала (Ес)
Таким образом, следует иметь в виду, что в случае постсинаптического торможения ТПСП не всегда имеют вид гиперполяризующих сигналов. Во многих случаях действие тормозного медиатора, вызывая увеличение хлорной проводимости мембраны, приводит к торможению нейрона путем шунтирования токов, входящих через возбуждающие синапсы нейрона, тем самым снижая или прекращая генерацию ВПСП и ПД.
Пресинаптическое торможение встречается в ЦНС наряду с постсинаптическим торможением. Примером является высвобождение тормозного медиатора ГАМК в аксо-аксоналыюм синапсе (рис. 3.34). Такой синапс образован окончанием аксона тормозного нейрона, выделяющего ГАМК, на нервной терминали другого синапса. ГАМК, выделяясь в таком аксо-аксональном синапсе, приводит к активации хлорных каналов своих рецепторов. При этом из-за высокого потенциала покоя мембраны аксона

Рис. 3.34. Пресинаптическое торможение на а-мотонейроне (а) и католическая депрессия в аксо-аксональном синапсе при длительной деполяризации его постсинаптической мембраны (б):
Сниженный по амплитуде ПД будет вызывать менее выраженную деполяризацию терминали, поэтому уменьшится Са 2+ -ток через потенциал- активируемые Са 2+ -каналы в активных зонах. Соответственно, снизится количество выбрасываемого медиатора, поскольку уровень экзоцитоза везикул критически зависит от поступления ионов Са 2+ в аксоплазму нервной терминали. В результате сниженной окажется и амплитуда ВПСП (рис. 3.35). Поскольку торможение нейрона происходит не на его постсинаптической мембране, а на пресинаптическом уровне, оно и называется пресинаптическим. Таким образом, пресинаптическое торможение имеет в своей основе предварительную деполяризацию мембраны аксона и тем самым ограничивает поступление полноценных нервных импульсов (ПД) к нервному окончанию нейрона.

Рис. 335. Пример пресинаптического торможения
Торможение играет важную роль в обработке поступающей в ЦНС информации. Особенно ярко эта функция выражена у пресинаптического торможения. К одному возбуждающему нейрону могут поступать сотни и тысячи сигналов по разным терминалям. Пресинаптическое торможение более точно регулирует процесс возбуждения, поскольку этим торможением могут быть заблокированы отдельные нервные волокна и синаптические входы нейрона. Число дошедших до нейрона импульсов определяется именно пресинап гическим торможением. В то же время постсинаптическое торможение обеспечивает более генерализованное снижение возбудимости нейрона.
Таким образом, и постсинаптическое, и пресинаптическое торможение со всеми их разновидностями выполняют охранительную роль для нейронов и синапсов. Отсутствие торможения приводило бы к истощению запаса медиаторов в синапсах и прекращению нормальной деятельности ЦНС. Блокада торможения ведет к широкой иррадиации возбуждения и судорогам. Такая ситуация наблюдается, например, при выключении пресинаптического ГАМКергического торможения в спинном мозге под действием блокатора ГАМК-рецепторов бикукулина.
Взаимодействие ВПСП и ТПСП на мембране постсинаптического нейрона приводит к флуктуациям мембранного потенциала, представляющего собой алгебраическую сумму деполяризующих и гиперполяризующих влияний. Так, на одном дендритном шипике могут присутствовать как тормозные, так и возбуждающие синапсы, т.е. будут практически одновременно возникать ТПСП и ВПСП. В этом случае конечный сдвиг потенциала будет представлять собой алгебраическую сумму ТПСП и ВПСП. Поэтому мембрана шипика, как и мембраны дендритов и тела нейрона, функционирует как определенного вида интегрирующее устройство, где происходит электрическое взаимодействие противоположно направленных электрических потенциалов.
На рис. 3.36 видно, что возбуждающие химические синапсы (показаны светлым) присутствуют в основном на дендригах нейрона. Эти синапсы весьма многочисленны и перемежаются с тормозными. Отдельные ВПСП никогда нс обеспечивают поодиночке достижение значения КУД. Тормозные синапсы (показаны темным) осуществляют постсинаптическое торможение. Они особенно многочисленны на соме нейрона, в районе аксонного холмика. Поэтому такие синапсы способны оказывать более сильное тормозное влияние на процессы нейронной интеграции.

Рис. 3.36. Расположение возбуждающих и тормозных входов на дендритах и соме нейрона
Тормозное влияние реализуется не только путем генерации гиперполя- ризующих ТПСП, но и за счет возникновения шунтирующей хлорной проводимости. При этом ГАМК открывает хлорные каналы ГАМК-рецепторов, но ток по ним нс идет, а за счет открывания CI -каналов происходит шунтирование возбуждающих Na + - и Са 2+ -токов. Так, в моторных нейронах кошки генерация одного ТПСП в районе аксонного холмика приводит к снижению Rax от 1—2 до 0,5 МОм. Это сопровождается снижением амплитуды суммарных ВПСП, достигающих аксонного холмика, в четыре и более раз (рис. 3.37).

Рис. 3.37. Соотношение ВПСП и ТПСП (и вызывающих их постсинаптических токов) при их одновременной генерации
Таким образом, нейрон способен интегрировать синаптические сигналы и выдавать на выходе в наиболее возбудимой части клетки — аксонном холмике — результирующий сигнал в виде ПД (рис. 3.38).

Рис. 3.38. Примеры пространственной и временной суммаций ВПСП и ТПСП как результат интеграции трех синаптических входов на нейроне (А и В — возбуждающие, С — тормозной)
Изучив принципы рефлекторной регуляции функций, процессы возникновения и проведения возбуждения, вы поняли, что на все эти процессы можно воздействовать - можно блокировать проведение импульса новокаином, можно увеличить или уменьшить возбудимость ткани, изменив ионный состав окружающей среды, можно воздействовать синаптоактивными веществами. Однако есть механизм, с помощью которого сам организм воздействует на процессы возбуждения и проведения импульса - торможение. Суть этого великолепного достижения эволюции необыкновенно проста: первый вариант - снижение возбудимости клетки вплоть до полной ее невозбудимости, второй вариант - изменение частоты идущих к клетке возбуждающих импульсов вплоть до прекращения проведения. Механизмы торможения - это преподнесенный нам в руки природой еще один путь воздействия на процессы возбуждения и проведения нервного импульса - можно воздействовать на процессы торможения, но для этого их нужно хорошо понять.
С л о в а р ь т е м ы:
Тормозные нейроны
Тормозные медиаторы
Пресинаптическое торможение, Постсинаптическое торможение
Первичное торможение, Вторичное торможение
Торможение Введенского (пессимальное)
Под ТОРМОЖЕНИЕМ понимают самостоятельный нервный активный процесс, вызываемый возбуждением, который проявляется в угнетении или полном выключении другого возбуждения. Торможение обусловлено нервными импульсами, возникающими в особых тормозных нейронах (клетки Рэншоу в спинном мозге, клетки Пуркинье коры мозжечка, звездчатые клетки коры головного мозга и др.).
Впервые возможность влияния на рефлекторные процессы без воздействия на структуры рефлекторной дуги была обнаружена И.М. Сеченовым. В 1863 году И.М. Сеченовым было открыто ЦЕНТРАЛЬНОЕ ТОРМОЖЕНИЕ (СЕЧЕНОВСКОЕ). И.М. Сеченову удалось установить, что сверхпороговое возбуждение центров среднего мозга приводит к торможению периферических, спинальных рефлексов.
Рисунок 23. Схема опыта И.М.Сеченова для демонстрации центрального торможения до и после аппликации на зрительные бугры NaCl.

Каким же образом можно изменить свойства нейронов так, чтобы они стали невозбудимы? Вспомним, в каких состояниях может находиться нейрон.
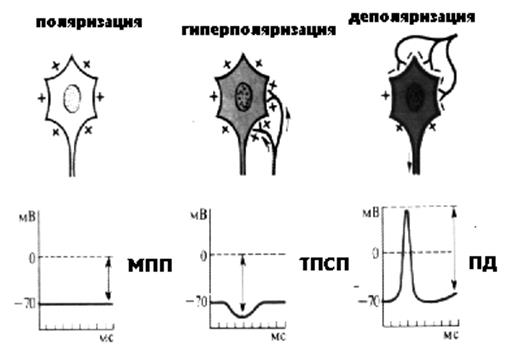
Рисунок 24 Состояния нейрона
Нас интересует торможение как самостоятельный процесс. ЦНС обладает механизмами изменения свойств нейрона без предварительного возбуждения. Вспомним результат выделения медиатора пресинаптической мембраной: на постсинаптической мембране происходит деполяризация и возникает локальный ответ. Тормозные нейроны синтезируют другие медиаторы - тормозные. Такими медиаторами могут быть глицин, гамма-амино-масляная кислота (ГАМК). Эти медиаторы, взаимодействуя со своими рецепторами, открывают другие каналы, например калиевые или хлорные. Что же происходит в этом случае с постсинаптической мембраной?
Если вспомнить о том, что ионы калия движутся из клетки, то становится ясно, что при активации калиевых каналов происходит гиперполяризация мембраны и снижение возбудимости нейрона. Тормозной медиатор, как и возбуждающий, активирует рецептор управляемые каналы, однако не натриевые, а калиевые. Эта активация вызывает увеличение выхода ионов калия наружу и ГИПЕРПОЛЯРИЗАЦИЮ мембраны. Изменение потенциала мембраны при гиперполяризации называется тормозной постсинаптический потенциал (ТПСП). Во многих тормозных синапсах аналогичный эффект имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. Результатом ТПСП является удаление МПП от критического уровня деполяризации возбуждение становится либо вовсе невозможным, либо для возбуждения требуется значительно большая сила раздражителя. Нервные импульсы, возникающие при возбуждении тормозных нейронов, вызывают гиперполяризацию постсинаптической мембраны и тормозной постсинаптический потенциал (ТПСП), это наиболее распространенный тип торможения в нервной системе, который называется ПОСТСИНАПТИЧЕСКИМ ТОРМОЖЕНИЕМ. Постсинаптическое торможение является первичным торможением – самостоятельный процесс, вызванный возбуждением тормозных нейронов.
Понятно, что в случае деполяризующего медиатора изменение потенциала мембраны будет называться возбуждающий постсинаптический потенциал (ВПСП).
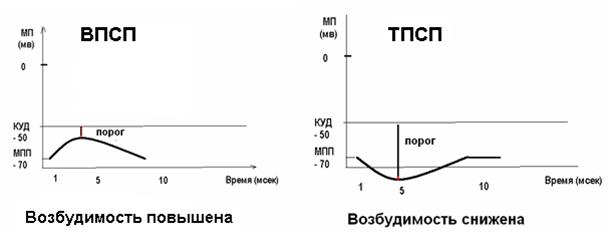
Рисунок 25 Изменение мембранного потенциала при ВПСП и ТПСП
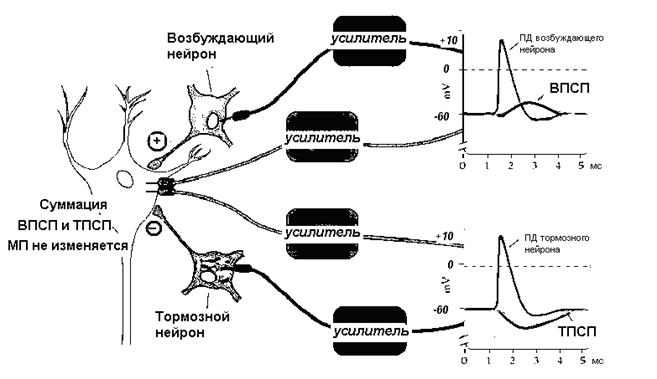
Рисунок 26 Суммация потенциалов на мембране нейрона
Центральные синапсы отличаются тем, что медиаторы вызывают появление на постсинаптической мембране только локального ответа, (ВПСП или ТПСП). ВПСП обладает всеми свойствами ЛО - зависит от силы раздражителя (количества выделившегося медиатора), суммируется и не передается по аксону. Амплитуда одиночного ВПСП довольно мала - несколько мВ, поэтому для уменьшения заряда мембраны до критического уровня необходима либо одновременная активация нескольких возбуждающих синапсов - ПРОСТРАНСТВЕННАЯ СУММАЦИЯ, либо повышение частоты поступающих импульсов - ВРЕМЕННАЯ СУММАЦИЯ. Если одновременно с ВПСП на мембране нейрона формируется и ТПСП, то эти изменения мембранного потенциала суммируются, следовательно, если величины ВПСП и ТПСП примерно одинаковы, то изменения мембранного потенциала нейрона, на котором суммируются воздействия, не произойдет. На рис.26 представлена схема регистрации мембранного потенциала возбуждающего и тормозного нейронов, которые одновременно воздействуют на один нейрон. Изменений мембранного потенциала нет. На рис 27 слева два возбуждающих нейрона, действуя на нейрон 1 одновременно, могли бы вызвать его возбуждение. Если одновременно на этот нейрон подействует тормозной, то возбуждения не произойдет.
Существует еще один вариант первичного торможения в ЦНС: ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ. Такое торможение возникает в том случае, когда нервный импульс или просто не достигает синаптического окончания аксона, или снижается его частота. Такое торможение осуществляется благодаря аксоаксональным синапсам, образованным терминалями аксонов тормозных интернейронов и аксонами возбуждающих нейронов (рис.27, справа). В этих синапсах торможение определяется действием ГАМК, открывающей хлорные каналы на пресинаптических мембранах (мембрана возбуждающего нейрона). Из - за высокого МПП мембраны аксона (действие электрического, а не концентрационного градиента) ионы хлора выходят через мембрану наружу. Выход ионов вызывает незначительную деполяризацию постсинаптической мембраны, являющейся в данном случае участком мембраны аксона другого нейрона. Деполяризация мембраны аксона ведет к инактивации натриевой проводимости. Возникает блокирование проведения нервного импульса, что уменьшает частоту, или полностью угнетает возбуждающий потенциал, идущий к клетке по возбуждающим нервным окончаниям.
| Постсинаптическое торможение | Пресинаптическое торможение |
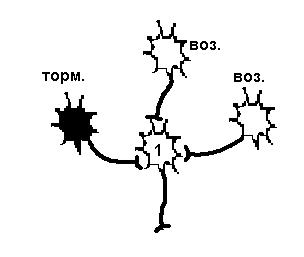 | 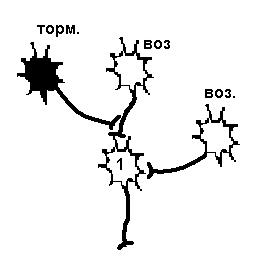 |
| Возбудимость нейрона 1 снижена в результате гиперполяризации мембраны, вызванной действием тормозного нейрона, поэтому два возбуждающих нейрона не могут вызвать ПД. | Возбудимость нейрона 1 не изменена, но возбуждающий импульс от одного из возбуждающих нейронов не доходит, а один возбуждающий не может вызвать ПД. |
Рисунок 27 Первичное торможение
| | | следующая лекция ==> | |
| Агонисты и антагонисты медиаторов | | | Распространение импульса в нейронных сетях |
Дата добавления: 2017-08-01 ; просмотров: 2184 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Медиатор, находящийся в пузырьках, выделяется в синаптическую щель с помощью экзоцитоза Его выделение происходит небольшими порциями – квантами. Небольшое количество квантов выходит из окончания и в состоянии покоя. Когда нервный импульс, т.е. ПД, достигает пресинаптического окончания, происходит деполяризация его пресинаптической мембраны. Открываются ее кальциевые каналы и ионы кальция входят в синаптическую бляшку. Начинается выделение большого количества квантов нейромедиатора. Молекулы медиатора диффундируют через синаптическую щель к постсинаптической мембране и взаимодействуют с её хеморецепторами. В результате образования комплексов медиатор-рецептор, в субсинаптической мембране начинается синтез так называемых вторичных посредников (в частности, цАМФ). Эти посредники активируют ионные каналы постсинаптической мембраны. Поэтому такие каналы называют хемозависимыми или рецепторуправляемыми. Т.е. они открываются при действии ФАВ на хеморецепторы. В результате открывания каналов изменяется потенциал субсинаптической мембраны. Такое изменение называется постсинаптическим потенциалом.
В ЦНС возбуждающими являются холин-, адрен-, дофамин-, серотонинергические синапсы и некоторые другие. При взаимодействии их медиаторов с соответствующими рецепторами, открываются хемозависимые натриевые каналы. Ионы натрия входят в клетку через субсинаптическую мембрану. Происходит ее местная или распространяющаяся деполяризация. Эта деполяризация называется возбуждающим постсинаптическим потенциалом (ВПСП).
Тормозными являются глицин- и ГАМК-ергические синапсы. При связывании медиатора с хеморецепторами, активируются калиевые или хлорные хемозависимые каналы. В результате ионы калия выходят из клетки через мембрану.
Величина ВПСП и ТПСП определяется количеством квантов медиатора, выделившихся из терминали, а, следовательно, частотой нервных импульсов. Т.е. синаптическая передача не подчиняется закону "все или ничего". Если количество выделившегося возбуждающего медиатора достаточно велико, то в субсинаптической мембране может генерироваться распространяющийся ПД. ТПСП, независимо от количества медиатора не распространяется за пределы субсинаптической мембраны.
ВОПРОС 26. Понятие о нервном центре, его функциях и свойствах
Н. центр – совокупность структур ЦНС, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт. Функциональный нервный центр может быть локализован в разных анатомических структурах. Например дыхат центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, коре головного мозга.
В зависимости от выполняемой функции различают:
чувствительные нервные центры;
нервные центры вегетативных функций;
двигательные нервные центры и др.
Свойства:
1)Одностороннее проведение возбуждения. В рефлекторной дуге, включающей н центры, процесс возбуждения распространяется в одном направлении (от входа по афферентным путям к центру, затем по эфферентным путям к эффектору).
2)Иррадиация возбуждения. В н центрах изменяется направление распространения возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов. Увеличение силы раздражителя приводит к расширению области вовлекаемых в возбуждение центральных нейронов – т. е. иррадиации возбуждения.
3)Суммация возбуждения. Процесс пространственной суммации афферентных потоков возбуждения от разл участков рецептивного поля облегчается благодаря наличию на мембране н клетки сотен и тысяч синаптичаских контактов. Процесс временной суммации в ответ на многократное возбуждение одних и тех же рецепторов обусловлены суммацией ВПСП на постсинаптической мембране.
ПОЯСНЯЮ: По́стсинапти́ческий потенциа́л (ПСП) — это вре́менное изменение потенциала постсинаптической мембраны в ответ на сигнал, поступивший с пресинаптического нейрона. Различают:
возбуждающий постсинаптический потенциал (ВПСП), обеспечивающий деполяризацию постсинаптической мембраны, и
тормозный постсинаптический потенциал (ТПСП), обеспечивающий гиперполяризацию постсинаптической мембраны.
Отдельные ПСП обычно невелики по амплитуде и не вызывают потенциалов действия в постсинаптической клетке, однако в отличие от потенциалов действия они градуальны и могут суммироваться. Выделяют два варианта суммации[1]:
временная — объединение пришедших по одному каналу сигналов (при поступлении нового импульса до затухания предшествующего)
пространственная — наложение ВПСП соседних синапсов
4) Наличие задержки.
5)Высокая утомляемость . Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до исчезновения. Это связано с деятельностью синапсов: истощение запаса медиатора, уменьшаются энергетических ресурсы, происходит адаптация постсинаптического рецептора к медиатору.
6)Тонус. В покое определенное количество нервных клеток находится в состоянии постоянного возбуждения и генерирует фоновые импульсные токи.
7)Пластичность. Функциональная подвижность нервного центра может модифицировать картину осуществляемых рефлекторных реакций.
8)Конвергенция. Нервные центры высших отделов мозга - мощные коллекторы афферентной информации. В них содержится много нервных клеток, реагирующих на разные стимулы ( свет, звук и др. )
9) Интеграция в нервных центрах. Для осуществления сложных координированных приспособительных реакций организма происходит образование функциональных объединений нервных центров.
10) Свойство доминанты. Доминантный очаг – временно господствующий очаг повышенной возбудимости в н центре. В нем устанавливается определенный уровень стационарного возбуждения, способствующий суммированию ранее подпороговых возбуждений и переводу на оптимальный для данных условий ритм работы. Домин. Очаг угнетает соседние очаги возбуждения.
11) Цефализация н. системы. Тенденция к перемещению функций регуляции и координации в головные отделы ЦНС.
ВОПРОС 27. Явление суммации возбуждения в нервных центрах, ее виды, значение, механизм. Свойства ВПСП и их роль в формировании суммации. ( Примечане автора: Ребят, я извеняюсь за эту хрень, но это все что я могла найти.
Суммация возбуждения. В работе нервных центров значительное место занимают процессы пространственной и временной суммации возбуждения, основным нервным субстратом которой является постсинаптическая мембрана. Процесс пространственной суммации афферентных потоков возбуждения облегчается наличием на мембране нервной клетки сотен и тысяч синаптических контактов. Процессы временной суммации обусловлены суммацией ВПСП на постсинаптической мембране.
В нервном волокне каждое одиночное раздражение (если оно не подпороговой и не свехпороговой силы) вызывает один импульс возбуждения. В нервных же центрах, как показал впервые И.М.Сеченов, одиночный импульс в афферентных волокнах обычно не вызывает возбуждения, т.е. не передается на эфферентные нейроны. Чтобы вызвать рефлекс необходимо быстрое нанесение допороговых раздражений одно за другим. Это явление получило название временной или последовательной суммации. Ее сущность состоит в следующем. Квант медиатора, выбрасываемого окончанием аксона при нанесении одного допорогового раздражения, слишком мал для того, чтобы вызвать возбуждающий постсинаптический потенциал, достаточный для критической деполяризации мембраны. Если же к одному и тому же синапсу идут быстро следующие один за другим допороговые импульсы, происходит суммирование квантов медиатора, и наконец его количество становится достаточным для возникновения возбуждающего постсинаптического потенциала, а затем и потенциала действия. Кроме суммации во времени, в нервных центрах возможна пространственнаясуммация. Она характеризуется тем, что если раздражать одно афферентное волокно раздражителем допороговой силы, то ответной реакции не будет, а если раздражать несколько афферентных волокон раздражителем той же допороговой силы, то возникает рефлекс, так как импульсы, приходящие с нескольких афферентных волокон суммируются в нервном центре.
Возбуждающий постсинаптический потенциал. В синапсах, в которых осуществляется возбуждение постсинаптической структуры, обычно происходит повышение проницаемости для ионов натрия. По градиенту концентрации Na+ входят в клетку, что вызывает деполяризацию постсинаптической мембраны. Эта деполяризация получила название: возбуждающий постсинаптический потенциал – ВПСП. ВПСП относится к локальным ответам и, следовательно, обладает способностью к суммации. Выделяют временную и пространственную суммацию.
Роль в суммации;
Принцип временной суммации - заключается в том, что импульсы поступают к пресинаптическому окончанию с периодом меньшим, чем период ВПСП.
Сущность пространственной суммации заключается в одновременной стимуляции постсинаптической мембраны синапсами, расположенными близко друг от друга. В этом случае ВПСП каждого синапса суммируются.
Если величина ВПСП достаточно велика и достигает критического уровня деполяризации (КУД), то генерируется ПД. Однако не все участки мембраны обладают одинаковой способностью к генерации ВПСП.
ВОПРОС 28.Явление трансформации ритма возбуждений в нервных центрах и его механизмы. Роль ВПСП и кольцевых связей в ЦНС. (Прим; Такая же херня что и с предыдущим вопросом -I’m sorryL)
Лат. transformatio - преобразование, превращение - одно из свойств проведения возбуждения в центре, заключающееся в способности нейрона изменять ритм приходящих импульсов. Особенно четко проявляется трансформация ритма возбуждения при раздражении афферентного волокна одиночными импульсами. На такой импульс нейрон отвечает серией импульсов. Это обусловлено возникновением длительного возбуждающего постсинаптического потенциала (роль ВПСП), на фоне которого развивается несколько ликов (спайков- пиковых потенциалов). Другой причиной возникновения множественного разряда импульсов являются следовые колебания мембранного потенциала. Когда его величина достаточно велика, следовые колебания могут привести к достижению критического уровня деполяризации мембраны и обусловливают появление вторичных спайков. В нервных центрах может происходить и трансформация силы импульсов: слабые импульсы усиливаются, а сильные ослабевают.
ВОПРОС 29. Посттетаническая потенциация в нервных центрах.(Тут мало – но это все что было в учебнике)
Это интегративный феномен. При раздражении афферентного нерва стимулами с низкой частотой можно получить рефлекс определенной интенсивности. Если затем этот нерв подвергать высокочастотному ритмическому раздражению, то повторное редкое ритмическое раздражение приведет к резкому усилению реакции.
ВОПРОС 30. Одностороннее проведение возбуждения в нервных центрах. Роль синаптических структур.
Одностороннее проведение возбуждения. В рефлекторной дуге, включающей н центры, процесс возбуждения распространяется в одном направлении (от входа по афферентным путям к центру, затем по эфферентным путям к эффектору).
Роль синаптических структур.
В отличие от нервных и мышечных волокон, для которых характерен закон двухстороннего проведения, в синапсе возбуждение распространяется только в одном направлении – от пресинаптической клетки к постсинаптической.
31.Высокая утомляемость нервных центров:
Утомление-ослабление рефлекторной реакции вплоть до ее полного исчезновения, происходящее под действием длительного повторного раздражения рецептивного поля рефлекса. Высокая утомляемость связана с деятельностью синапсов, в которых запасы медиатора истощаются ,уменьшаются энергетические ресурсы. а также высокая утомляемость нервных центров происходит из-за адаптации постсинаптических рецепторов к медиаторам.
32.тонус нервных центров и его механизмы:
Тонус-наличие определённой фоновой активности нервного центра. То есть ,в покое, в отсутствие внешних раздражителей определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки . например, во сне в высших отделах мозга остаётся некоторое количество фоновоактивных нервных клеток, определяющих тонус соответствующего нервного центра.
Читайте также:
